











Van der Made Jan
- Investigador Científico
- Departamento: Paleobiología
- Teléfono: 915668966 ext. 988966
- Email: jvdm@mncn.csic.es
- Página web externa: https://scholar.google.com/citations?user=0mcYuFUAAAAJ&hl=en
- Número ORCID: 0000-0002-4807-4338
- Palabras clave:

Investigación
Initially, I mainly studied the Suoidea (= pigs and peccaries) from Europe, Asia and Africa, and from the Eocene till recent, but also the contemporary ungulates, primarily Bovidae, Cervidae, Tragulidae and Anthracotheriidae. Since 1993, I am the coordinator for the Suoidea of the NOW database.
When I became a member of the Atapuerca team, my research shifted to the ungulates of the Pleistocene. This is also a shift in the approach, since prehistoric research is centred on particular sites which are often excavated during many years. With the idea of synergy, I take or took part in numerous other projects on prehistoric sites, foremost the Algerian sites of Ain Hanech, Ain Boucherit, El Kherba and Tighenif, but also Bilzingsleben and Neumark Nord (Germany), Khoramabad Valley (Iran), Armenia, Morocco, etc.
I am much interested in the effects of climatic and geographic change on the fauna. A special case is insular environment. I was a student of P.Y. Sondaar, who was a specialist in insular ecology and evolution, and belong to his “school”.
In the following sections, my papers are listed which are dedicated to or contribute to the different categories. The same paper may appear in more than one section. Most of the links are to websites of publishers. If you do not have access, please write me an e-mail.
1) Research by sites: palaeontology and the relationship with human prehistory
2) Evolution, systematics and biogeography of Suoidea and other groups
3) Biogeography, biostratigraphy, "the history of nature", extinction
4) Insular ecology and evolution
5) Others
1) Research by sites: palaeontology and the relationship with human prehistory
Atapuerca
Since 1995, I participate in the Atapuerca project, where it is my task to study the ungulates.
The main sites from which I studied the ungulates are Gran Dolina, Galería and Sima del Elefante. There are but few fossils from Penal and Cueva del Fantasma is a new site. Sima de los Huesos is an important site, but does not have ungulates. The open air sites (Valle de las Orquídeas, El Hundidero, Hotel California and Fuente Mudarra) do not have fauna. Other sites, like El Mirador, have mainly Holocene and the fauna is mainly domestic.
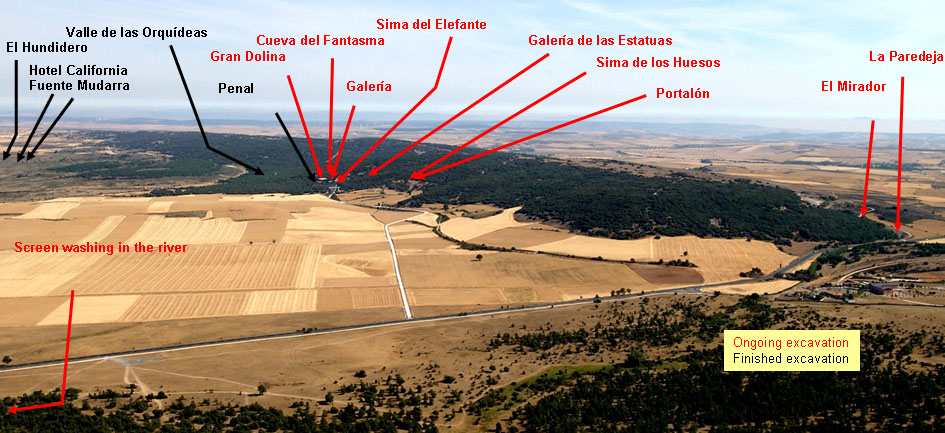
The different localities in the Sierra de Atapuerca.
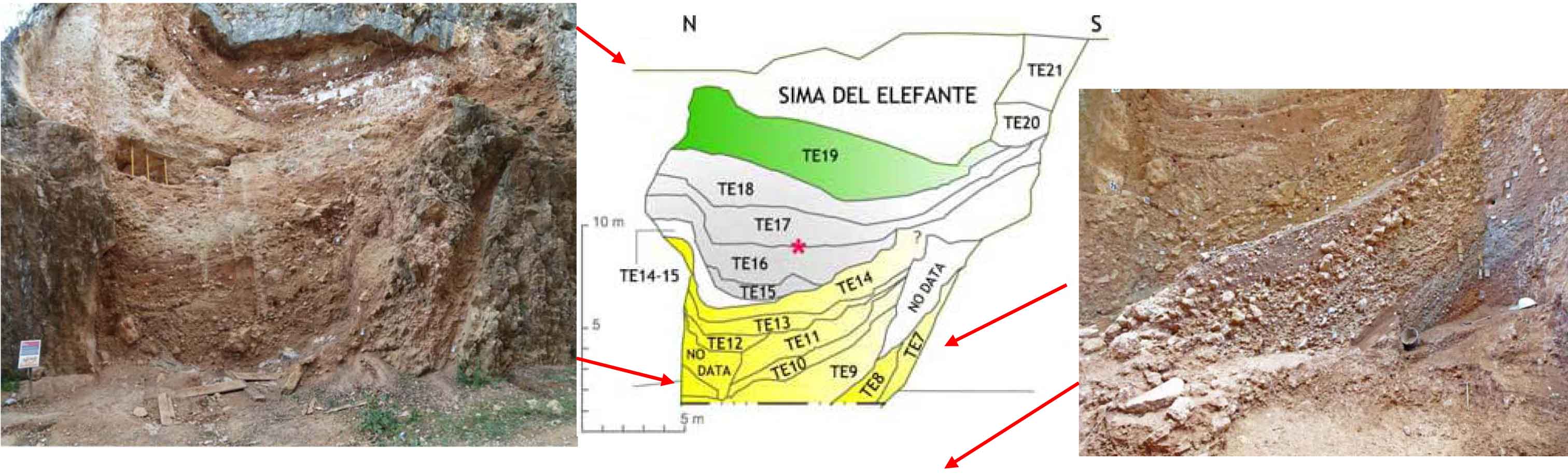
Mosaic photograph of Sima del Elefante in 2006 before excavation below the base level of the railway trench (left), stratigraphic scheme of the upper part (middle), and excavated part below the base level of the railway trench in 2013 (right).
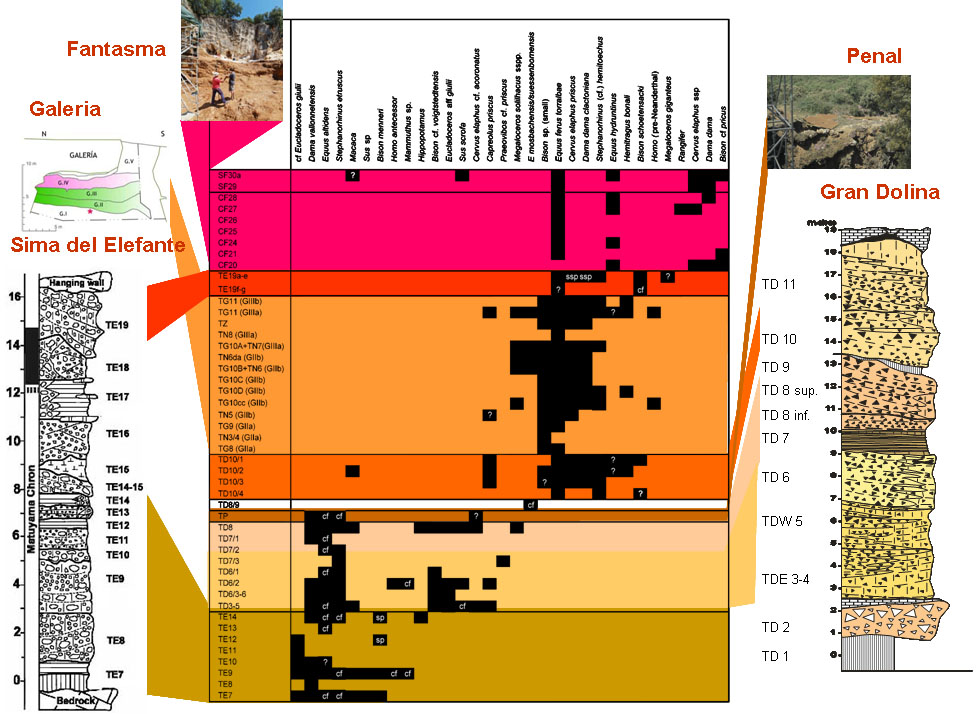
The distribution of the ungulate species in the different sites and levels at Atapuerca (adapted from Van der Made, 2013).
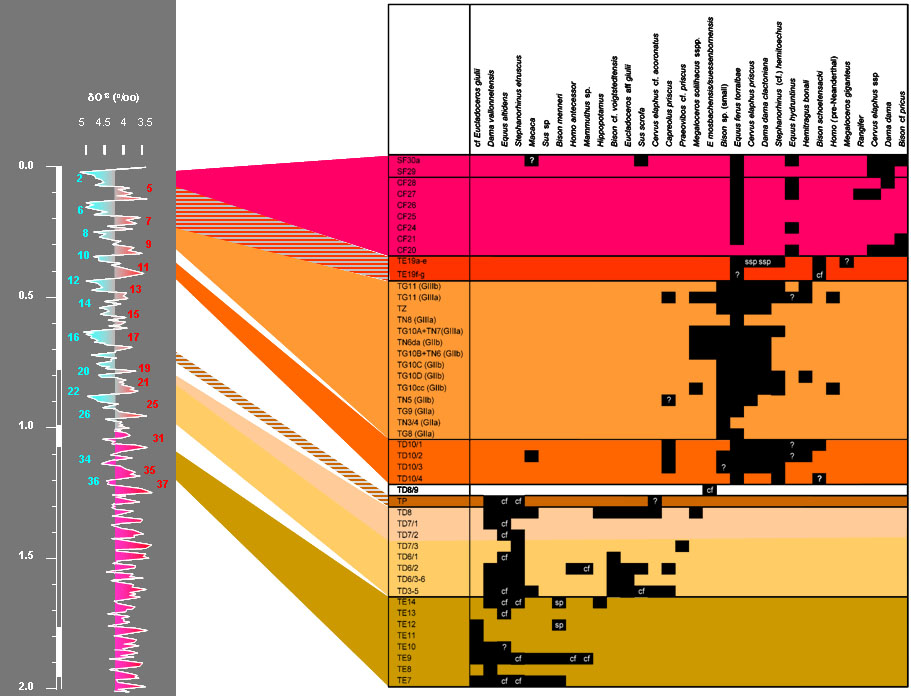
The temporal distribution of the ungulates from the different sites and levels at Atapuerca (adapted from Van der Made, 2013).
-
Made, J. van der , I.A. Lazagabaster, P. García-Medrano & I. Cáceres, 2026. Southernmost Eurasian Record of Reindeer (Rangifer) in MIS 8 at Galería (Atapuerca, Spain): Evidence of Progressive Southern Expansion of Glacial Fauna Across Climatic Cycles. Quaternary, 8, 43.
-
-
Rodríguez-Gomez, G., A. Rodríguez-Hidalgo, P. Saladié, J. van der Made, J. Marín, A. Ollé, M. Mosquera, J. Maria Bermúdez de Castro, J.L. Arsuaga & E. Carbonell, 2025. Ecologically sustainable human exploitation of the Gran Dolina TD10.2 bison (Sierra de Atapuerca, Spain). Scientific Reports, 15:23178.
-
-
Huguet, R., X.P. Rodríguez-Álvarez, M. Martinón-Torres, J. Vallverdú, J.M. López-García, M. Lozano, M. Terradillos-Bernal, I. Expósito, A. Ollé, E. Santos, P. Saladié, A. de Lombera-Hermida, E. Moreno-Ribas, L. Martín-Francés, E. Allué, C. Núñez-Lahuerta, J. van der Made, J. Galán1, H.A. Blain, I. Cáceres, A. Rodríguez-Hidalgo, A. Bargalló, M. Mosquera, J.M. Parés, J. Marín, A. Pineda, D. Lordkipanidze, A. Margveslashvili, J.L. Arsuaga, E. Carbonell & J.M. Bermúdez de Castro, 2025. The earliest human face of Western Europe. Nature, 640: 707-713.
-
-
Made, J. van der, 2025. Did human dispersal into Europe cause the continent-wide extinction of the pig Sus strozzii at 1.8 Ma?—Review of a Debate. Quaternary, 8/26.
-
-
Álvarez-Posada, C., J.M. Parés, G. Cuenca-Bescós, J. van der Made, J. Rosell, J.M. Bermúdez de Castro & E. Carbonell, 2018. A post-Jaramillo age for the artefact-bearing layer TD4 (Gran Dolina, 1 Atapuerca): New paleomagnetic evidence. Quaternary Geochronolog, 45: 1-8.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.

Reconstruction of the landscape of Atapuerca TD8 in autumn (brown leaves) with Hippopotamus (today a tropical animal), Stephanorhinus etruscus (a rhinoceros) and Dama vallonnetensis (a primitive species of fallow deer) by Mauricio Antón (Bermúdez de Castro et al., 1999).
- Rodríguez-Gomez, G., A. Rodríguez-Hidalgo, P. Saladié, J. van der Made, J. Marín, A. Ollé, M. Mosquera, J. Maria Bermúdez de Castro, J.L. Arsuaga & E. Carbonell, 2025. Ecologically sustainable human exploitation of the Gran Dolina TD10.2 bison (Sierra de Atapuerca, Spain). Scientific Reports, 15:23178
- Huguet, R., J. Vallverdú, X.P. Rodríguez- Alvarez, M. Terradillos-Bernal, A. Bargalló, A. Lombera-Hermida, I. Menéndez, M. Modesto-Mata, J. van der Made, M. Soto, H.A. Blain, N. García, G. Cuenca-Bescós, G. Gómez-Merino, R. Pérez-Martínez, I. Expósito, E. Allué, J. Rofes, F. Burjachs, A. Canals, M. Bennásar, C. Nuñez-Lahuerta J.M. Bermúdez de Castro & E. Carbonell, 2017. Level TE9c of Sima del Elefante (Sierra de Atapuerca, Spain): A comprehensive approach. Quaternary International, 433: 278-295.
- Carbonell, E., R. Huguet, I. Cáceres, C. Lorenzo, M. Mosquera, A. Ollé, X.P. Rodríguez, P. Saladié, J.M. Vergès, P. García-Medrano, J. Rosell, J. Vallverdú, J.M. Carretero, M. Navazo, A.I. Ortega, M. Martinón-Torres, J.I. Morales, E. Allué, A. Aramburu, A. Canals, A. Carrancho, M. Castilla, I. Expósito, M. Fontanals, M. Francés, M. Galindo-Pellicena, D. García-Antón, N. García, A.Gracia, R. García, G. Gómez-Merino, E. Iriarte, A. Lombera-Hermida, L. López-Polín, M. Lozano, J. van der Made, I. Martínez, A. Mateos, A. Pérez-Romero, E. Poza, R. Quam, A. Rodriguez-Hidalgo, J. Rodríguez, L. Rodríguez, E. Santos, M. Terradillos, J.M. Bermúdez de Castro & J.L. Arsuaga, 2014. Los yacimientos arqueológicos de la Sierra de Atapuerca. In: R. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro & J.L. Arsuaga (eds). Los cazadores recolectores del Pleistoceno y del Holoceno en Iberia y el Estrecho de Gibraltar: estado actual del conocimiento del registro arqueológico. Universidad de Burgos & Fundación Atapuerca - Burgos: 534-560.
- Carbonell, E., R. Huguet, I. Cáceres, C. Lorenzo, M. Mosquera, A. Ollé, X.P. Rodríguez, P. Saladié, J.M. Vergès, P. García-Medrano, J. Rosell, J. Vallverdú, J.M. Carretero, M. Navazo, A.I. Ortega, M. Martinón-Torres, J.I. Morales, E. Allué, A. Aramburu, A. Canals, A. Carrancho, M. Castilla, I. Expósito, M. Fontanals, M. Francés, M. Galindo-Pellicena, D. García-Antón, N. García, A.Gracia, R. García, G. Gómez-Merino, E. Iriarte, A. Lombera-Hermida, L. López-Polín, M. Lozano, J. van der Made, I. Martínez, A. Mateos, A. Pérez-Romero, E. Poza, R. Quam, A. Rodriguez-Hidalgo, J. Rodríguez, L. Rodríguez, E. Santos, M. Terradillos, J.M. Bermúdez de Castro & J.L. Arsuaga, 2014. Sierra de Atapuerca archaeological sites. In: R. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro & J.L. Arsuaga (eds). Pleistocene and Holocene hunter-gatherers in Iberia and the Gibraltar Strait: the current archaeological record. Universidad de Burgos & Fundación Atapuerca - Burgos: 534-560.
- Made, J. van der, 2013. First description of the large mammals from the locality of Penal, and updated faunal lists for the Atapuerca ungulates - Equus altidens, Bison and human dispersal into Western Europe. Quaternary International, 295: 36-47.
- Made, J. van der, 2013. Clima, fauna y dispersión humana desde África a Eurasia. Periodico de Atapuerca, 25: 2.
- Made, J. van der, 2011. Biogeography and climatic change as a context to human dispersal out of Africa and within Eurasia. Quaternary Science Reviews, 30: 1353-1367.
- Rodríguez, J., F. Burjachs, G. Cuenca-Bescós, N. García, J. van der Made, A. Perez-González, H. A. Blain, I. Expósito, J.M. López-García, M. García-Antón, E. Allué, I. Cáceres, R. Huguet, M. Mosquera, A. Ollé, J. Rosell, J.M. Parés, X.P. Rodríguez, J.C. Díez, J. Rofes, R. Sala, P. Saladié, J. Vallverdú, M.L. Bennasar, R. Blasco, E. Carbonell, J.M. Bermúdez de Castro, 2011. One million years of cultural evolution in a stable environment at Atapuerca (Burgos, Spain). Quaternary Science Reviews, 30: 1396-1412. Doi:10.1016/j.quascirev.2010.02.021.
- Blasco, R., J. Rosell, J. van der Made, J. Rodríguez, G. Campeny, J.L. Arsuaga, J.M. Bermúdez de Castro & E. Carbonell, 2011. Hiding to eat: the role of carnivores in the early Middle Pleistocene from the TD8 level of Gran Dolina (Sierra de Atapuerca, Burgos, Spain). Journal of Archaeological Science, 38: 3373-3386.
- Made, J. van der, 2010. The ungulates from the Atapuerca sequence and large mammal dispersals from Asia into Europe. Communication on Contemporary Anthropology: 12(9): 143-145.
- Made, J. van der & A. Mateos, 2010. Longstanding biogeographic patterns and the dispersal of early Homo out of Africa and into Europe. Quaternary International, 223-224: 195-200.
- Rosell, J., R. Blasco, J. van der Made, I. Cáceres, G. Campeny, J.C. Díez, P. Saladié, E. Carbonell, 2010. Acumulaciones producidas por carnívoros en el Pleistoceno Medio inicial: una reconstrucción a partir del conjunto faunístico del nivel TD8 de Gran Dolina (Sierra de Atapuerca, Burgos). Zona Arqueológica, 13: 158-171.
- Carbonell, E., J.M. Bermúdez de Castro, J.M. Parés, A. Pérez-González, A. Ollé, M. Mosquera, G. Cuenca-Bescós, N. García, D.E. Granger, R. Huguet, J. van der Made, M. Martinón-Torres, X.P. Rodríguez, A. Rosas, R. Sala, G.M. Stock, J. Vallverdú, J.M. Vergès, E. Allué, F. Burjachs, I. Cáceres, A. Canals, A. Benito, C. Díez, M. Lozano, A. Mateos, M. Navazo, J. Rodríguez, J. Rosell, J.L. Arsuaga, 2008. The first hominin of Europe. Nature, 452:465-469. Supplementary information.
- Made, J. van der, 2008. La Sima del Elefante proporciona el resto humano más antiguo de Europa. Periódico del Museo Nacional de Ciencias Naturales, 6 (abril-junio 2008): 12.
- Rosas, A., R. Huguet, A. Pérez-González, E. Carbonell, J. M. Bermúdez de Castro, J. Vallverdú, J. van der Made, E. Allué, N. García, R. Martínez-Pérez, J. Rodríguez, R. Sala, P. Saladie, A. Benito, C. Martínez-Maza, M. Bastir, A. Sánchez, J. M. Parés, 2006. The “Sima del Elefante” cave site at Atapuerca (Spain). Estudios Geológicos, 62 (1): 327-348.
- Carbonell, E., J.M. Bermúdez de Castro, J.L. Arsuaga, E. Allue, M. Bastir, A. Benito, I. Cáceres, T. Canals, J.C. Díez, J. van der Made, M. Mosquera, A. Ollé, A. Pérez-González, J. Rodríguez, X.P. Rodríguez, A. Rosas, J. Rosell, R. Sala, J. Vallverdú & J.M. Vergés, 2005. An Early Pleistocene hominin mandible from Atapuerca-TD6, Spain. Proceedings of the National Academy of Sciences, 102(16): 5674-5678.
- Made, J. van der, 2005. Considerations on dispersals between Africa and Europe across the Strait of Gibraltar. In: J. Rodríguez Vidal,c. Finlayson & F. Giles Pacheco (eds.). Cuaternario Mediterraneo y poblamiento de hominidos. AEQUA & Gibraltar Museum, Gibraltar: 91-92.
- Bermúdez de Castro, J.M., M. Martinón-Torres, E. Carbonell, S. Samiento, A. Rosas, J. van der Made & M. Lozano, 2004. The Atapuerca sites and their contribution to the knowledge of human evolution in Europe. Evolutionary Anthropology, 13: 25-41.
- Made, J. van der, 2004. La paleontología, ¿para qué sirve? Diario de los yacimientos de la Sierra de Atapuerca, 11: 1-2. (supplemento del Diario de Burgos)
- Rosas, A., R. Huguet, A. Pérez-González., E. Carbonell, J. Vallverdú, J. van der Made; E. Allué, N. García, R. Martínez-Pérez, J. Rodríguez, R. Sala, P. Saladie, G. Simón, C. Martínez-Maza, M. Bastir, A. Sánchez & J.M. Parés, 2004. Initial approach to the site formation and paleoecology of the “Sima del Elefante”: a Pleistocene karst site at Atapuerca Hill. Zona Arqueologica, 4(1): 134-155.
- Cuenca Bescós, G., N. García & J. van der Made, 2004. Fossil mamals of the Lower Pleisotcene site of Gran Dolina, Atapuerca (Burgos, Spain). Zona Arqueologica, 4(4): 140-149.
- Rosas, A., A. Pérez-González, E. Carbonell, J. van der Made, A. Sánchez, C. Laplana, G. Cuenca-Bescós, J.M. Parés & R. Huguet, 2001. Le gisement pléistocène de la "Sima del Elefante" (Sierra de Atapuerca, Espagne). L'Anthropologie, 105 (2): 301-312.
- Made, J. van der, 2001. Les ongulés d'Atapuerca. Stratigraphie et biogéographie. L'Anthropologie, 105 (1): 95-113.
- Made, J. van der, 2001. Biogeografía y colonización de Europa. La Vanguardia, lunes, 5 de noviembre, 2001: p. 29.
- Arsuaga, J.L., J.M. Bermúdez de Castro, E. Carbonell & J.J. Fernández (eds.), E. Allué, B. Arnaíz, J.L. Arsuaga, J.M. Bermúdez de Castro, I. Cáceres, J.I. Canudo, E. Carbonell, J.M. Carretero, G. Cuenca, J.C. Díez, A. Esquivel, J.J. Fernández, Y. Fernández-Jalvo, N. García, D. García-Antón, A. Gracia, R. Huguet, N. Ibañez, C. Laplana, L. López-Polín, C. Lorenzo, J. van der Made, C. Mallol, B. Márquez, A. Martín, I. Martínez, M. Mosquera, M.E. Nicolás, A. Ollé, A.I. Ortega, J.M. Parés, A. Pérez-González, P.J. Pérez, A. Pérez-Pérez, J. Rodríguez, X.P. Rodríguez, A. Rosas, J. Rosell, R. Sala, P. Saladié, A. Sánchez, S. Sarmiento, J. Vallerdú, J.M. Vergés, 2000. Los yacimientos de la Sierra de Atapuerca, Burgos. Guía Arqueológica. Fundación del Patrimonio Histórico de Castilla y León: 1-73.
- Bermúdez de Castro, J.M., A. Rosas, J. van der Made & J. Rodriguez, 2000. Human fossil remains from the Gran Dolina Lower Pleistocene site (Siera de Atapuerca, Spain). Études et Recherches Archéologiques de l'Université de Liège, 92: 161.
- Rodriguez, J., J. van der Made, A. Rosas & J.M. Bermúdez de Castro, 2000. Fauna and Palaeoecology of Gran Dolina-TD6 (Sierra de Atapuerca, Spain). Études et Recherches Archéologiques de l'Université de Liège, 92: 163
- Bermúdez de Castro, J.M., J.L. Arsuaga, E. Carbonell & J. Rodríguez (eds.), J.J. Fernández Moreno, E. Aguirre, E. Allué, J.L. Arsuaga, J.M. Bermúdez de Castro, I. Cáceres, J.I. Canudo, E. Carbonell, J.M. Carretero, G. Cuenca, J.C. Díez, A. Esquivel, Y. Fernández-Jalvo, N. García, D. García-Antón, A. Gracia, R. Huguet, C. Laplana, L. López-Polín, C. Lorenzo, J. van der Made, C. Mallol, B. Marquéz, A. Martín, I. Martínez, M. Mosquera, M.E. Nicolás, A. Ollé, A.I. Ortega, J.M. Parés, A. Pérez-González, P.J. Pérez, A. Pérez-Pérez, J. Rodríguez, A. Rosas, J. Rosell, R. Sala, A. Sánchez, S. Sarmiento, J. Vallerdú, J.M. Vergés, C. Cubillo, M. Antón, 1999. Atapuerca - Nuestros antecessores. Junta de Castilla y León: 1-221.
- Bermúdez de Castro, J.M., E. Carbonell, I. Cáceres, J.C. Díez, Y. Fernández-Jalvo, M. Mosquera, A. Ollé, J. Rodríguez, A. Rosas, J. Rosell, R. Sala, J.M. Vergés & J. van der Made, 1999. The TD6 (Aurora Stratum) hominid site. Final remarks and new questions. Journal of Human Evolution, 37(3-4): 695-700.
- Carbonell, E., M. Mosquera, X.P. Rodríguez, R. Sala & J. van der Made, 1999. Out of Africa: the Dispersal of the Earliest Technical Systems Reconsidered. Journal of Anthropological Archaeology, 18: 119-136.

Reconstruction of Atapuerca TD6 by Mauricio Antón with Eucladoceros aff. giulii (a large species of deer), the bison Bison voigtstedtensis and the oldest Sus scrofa (wild boar) of Europe (Bermúdez de Castro et al., 1999).
- Made, J. van der, 1999. Ungulates from Atapuerca-TD6. Journal of Human Evolution, 37(3-4): 389-413.
- Made, J. van der, 1999. Artiodactylos del yacimiento mesopleistoceno de Galería (Sierra de Atapuerca). En E. Carbonell, A. Rosas & C. Diez (eds.): Ocupaciones humanas y paleoecología del yacimiento de Galería. Junta de Castilla y León: 143-167.
- Rosas, A., E. Carbonell, A. Ollé, A. Pérez González, J. Vallverdú, R. Huguet, I.Cácers, J. Rosell, R. Sala, M. Mosquera, J. van der Made, A. Sánchez, G. Cuenca, X.P. Rodrígquez & J. Rodríguez, 1999. Contribución del yacimiento de Galería (Sierra de Atapuerca) al Cuaternario Ibérico. En E. Carbonell, A. Rosas & C. Diez (eds.): Ocupaciones humanas y paleoecología del yacimiento de Galería. Junta de Castilla y León: 377-390.
- Made, J. van der, 1998. Ungulates from Gran Dolina (Atapuerca, Burgos, Spain). Quaternaire, 9(4): 267-281.
- Rosas, A., E. Carbonell, G. Cuenca, N. García, Y. Fernández Jalvo, J. van der Made, M. Ollé, J.M. Parés, A. Pérez González, A. Sánchez Marco, B. Sánchez Chillón & J. Valverdú, 1998. Cronología, bioestratigrafía y paleoecología del Pleistoceno Medio de Galería (Sierra de Atapuerca). Revista Española de Paleontología, 13(1): 71-80.
- Rosas, A., J.M. Bermúdez de Castro & J. van der Made, 1997. Homo antecessor en zijn paleontologische context in Gran Dolina (Atapuerca). Cranium, 14(2): 71-77.
Algeria: Ain Boucherit, Ain Hanech, El Kherba & Tighennif
Since 2002, I participate in a project, lead by Mohamed Sahnouni, on localities near El Eulma and later also Tighennif. The localities Ain Boucherit, Ain Hanech, El Kherba and others are in a small valley, the Oued Boucherit and form a magnetostratigraphically datedsequence covering the time from >3 Ma to <1.7 Ma. Tighennif is a younger, late Early Pleistocene locality. The main objective of the project is prehistory and my task is the study of the large mammals. One of our most relevant results is the excavation of stone tools in Ain Boucherit, which date to 2.44 Ma.
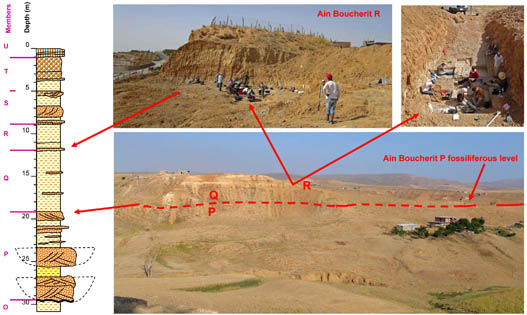
Ain Boucherit: the fossiliferous level at the top of member P and the excavation in member R of the Ain Hanech Fm.

El Kherba with Ain Boucherit in the background. Photograph by Jordi Mestre.
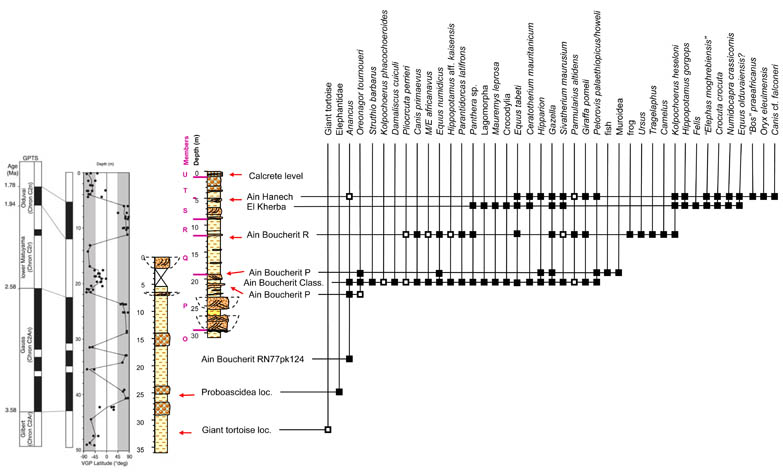
Biostratigraphy of the localities in the Oued Boucherit (Van der Made et al., 2021).
- Armaroli, E., R. Chelli Cheheb, A. Cipriani, S. Bernardini, J. van der Made, I. Cáceres, M. Sahnouni & F. Lugli, 2025. Stable Sr isotopes of fossil dental enamel reflect diet and digestive system differences among sympatric herbivores. Palaeogeography, Palaeoclimatology, Palaeoecology, 678: 113226.
- Cáceres, I., R. Chelli Cheheb, J. van der Made, Z. Harichane, K. Boulaghraief & M. Sahnouni, 2023. Assessing the subsistence strategies of the earliest North African inhabitants: evidence from the Early Pleistocene site of Ain Boucherit (Algeria). Archaeological and Anthropological Sciences, 15:87.
- Duval, M., M. Sahnouni, J.M. Parés, J.-x. Zhao, R. Grün, S. Abdessadok, A. Pérez-González, A. Derradji, Z. Harichane, N. Mazouni, K. Boulaghraief, R. Chelli Cheheb, J. van der Made, 2023. On the age of Ain Hanech Oldowan locality (Algeria): First numerical dating results. Journal of Human Evolution, 180: 103371.
- Made, J. van der Made, K. Boulaghraief, R. Chelli- Cheheb, I. Cáceres, Z. Harichane & M. Sahnouni, 2022. The last North African hipparions – hipparion decline and extinction follows a common pattern. Neues Jahrbuch fürGeologie und.Paläontologie, Abhandlungen, 303/1: 39-87.
- Duval, M., M. Sahnouni, J.M. Parés, J. van der Made, S. Abdessadok, Z. Harichane, R. Chelli Cheheb, K. Boulaghraif, A. Pérez-González, 2021. The Plio-Pleistocene sequence of Oued Boucherit (Algeria): A unique chronologically-constrained archaeological and palaeontological record in North Africa. Quaternary Science Reviews, 271: 107116.
- Made, J. van der, K. Boulaghraief, R. Chelli-Cheheb, M. Duval, J.M. Pares, I. Caceres, S. Semaw & M. Sahnouni, 2021. The North African earliest Pleistocene faunal sequence: from biostratigraphy to biochronology. Mémoire du Service Géologique de l’Algérie, 22: 73-90.
- Sahnouni Mohamed, J.M. Parés, M. Duval, I. Cáceres, Z. Harichane, J. van der Made, A. Pérez-González, S. Abdessadok, N. Kandi, A. Derradji, M. Medig, K. Boulaghraif, R. Chelli Cheheb, S. Semaw, 2021. Mise en évidence d’outils lithiques et de traces de découpe datés de 2,44 et de 1,92 millions d’années dans le site de Aïn Boucherit (Sétif, Algérie) et leurs implications pour la première occupation humaine en Afrique du Nord. Mémoire du Service Géologique de l’Algérie, 22: 55-72.
- Sahnouni, M., J.M. Parés, M. Duval, I. Cáceres, Z. Harichane, J. van der Made, A. Pérez-González, S. Abdessadok, N. Kandi, A. Derradji, M. Medig, K. Boulaghraif & S. Semaw, 2018. 1.9-million- and 2.4-million-year-old artifacts and stone tool–cutmarked bones from Ain Boucherit, Algeria. Science, 362, 1297–1301. Supplementary Materials.
- Cáceres, I., N. Kandi, M. Sahnouni, Z. Harichane & J. van der Made, 2017. Taphonomy of the Early Pleistocene fossil assemblages from Ain Boucherit (Ain Hanech, Algeria). In: M. Sahnouni, S. Semaw & J. Rios Garaizar (eds.) Proceedings of the II Meeting of African Prehistory, Burgos 15-16 April, 2015. Centro Nacinonal de Investigación sobre la Evolución Humana - Burgos: 171-196.
- Made, J. van der, M. Sahnouni & Kamel Boulaghraief, 2017. Hippopotamus gorgops from El Kherba (Algeria) and the context of its biogeography. In: M. Sahnouni, S. Semaw & J. Rios Garaizar (eds.) Proceedings of the II Meeting of African Prehistory, Burgos 15-16 April, 2015. Centro Nacinonal de Investigación sobre la Evolución Humana - Burgos: 135-169.
- Sahnouni, M., M. Everet, J. van der Made, Z. Harichane, 2017. Mise enévidence d’un changement climatique dans le site pléistocène inférieur d’El Kherba (Algérie), et son possible impact sur les activités des hominidés,il-y-a1,7Ma. L’Anthropologie, 121: 146-162.
- Sahnouni, M., A. Pérez-González, J. Parés, S. Abdessadok, J. van der Made, M. Duval, Z. Harichane, I. Cáceres, N. Kandi, K. Boulaghraief, A. Derradji, M. Medig, Y. Mouhoubi, A. Hamlat, S. Chibane, 2017. La primera ocupación humana en el norte de Africa: Evidencia de la secuencia plio-pleistocena de Ain Boucherit-Ain Hanech (Argelia). In: M. Sahnouni, S. Semaw & J. Rios Garaizar (eds.) Proceedings of the II Meeting of African Prehistory, Burgos 15-16 April, 2015. Centro Nacinonal de Investigación sobre la Evolución Humana - Burgos: 263-295.
- Parés, J.M., M. Sahnouni, J. van der Made, A. Pérez-González, Z. Harichane, A. Derradji & M. Medig, 2014. Early human settlements in Northern Africa: paleomagnetic evidence from the Ain Hanech Formation (northeastern Algeria). Quaternary Science Reviews, 99(1): 203–209.
- Made, J. van der, 2013. Faunal exchanges through the Levantine Corridor and human dispersal: The paradox of the latedispersal of the Acheulean industry. In: M. Sahnouni (ed.) Proceedings of the International Symposium Africa, Cradle Of Humanity: Recent Discoveries. Travaux du Centre National de Recherches Préhistoriques, Anthropologiques et Historiques, Nouvelle série 18: 255-296.
- Made, J. van der & M. Sahnouni, 2013. Updated Plio-Pleistocene faunal lists for Ain Boucherit, Ain Hanech, and El Kherba sites, Algeria. In: M. Sahnouni (ed.) Proceedings of the International Symposium Africa, Cradle Of Humanity: Recent Discoveries. Travaux du Centre National de Recherches Préhistoriques, Anthropologiques et Historiques, Nouvelle série 18: 223-242.
- Sahnouni, M., J. de Heinzelin, J. van der Made, F. Brown, M. Everett, Z. Harichane, N. Kandi, J. Rosell, D. Hadjouis, A. Derradji, A. Ollé, J.M. Vergès, M. Medig & T. Canals, 2013. La séquence plio-pléistocène d'Ain Boucherit-Ain Hanech (Algérie orientale): Biochronologie, environnements, et comportements hominidés. In: M. Sahnouni (ed.) Proceedings of the International Symposium Africa, Cradle Of Humanity: Recent Discoveries. Travaux du Centre National de Recherches Préhistoriques, Anthropologiques et Historiques, Nouvelle série 18: 189-210.
- Sahnouni, M., J. Rosell, J. van der Made, J.M. Vergès, A. Ollé, A. Derradji, Z. Harichane, N. Kandi, N. Mellah, M. Medig, 2013. The first evidence of cut marks and usewear traces from the Plio-Pleistocene locality of El-Kherba (Ain Hanech), Algeria. Implications for early hominin subsistence activities circa 1.8 Ma. Journal of Human Evolution, 64: 137-150.
- Sahnouni, M., J. van der Made & M. Everett, 2011. Ecological Background to Human occupation in North Africa during the Early Pleistocene Ecological background to Plio-Pleistocene hominin occupation in North Africa: the vertebrate faunas from Ain Boucherit, Ain Hanech and El-Kherba, and paleosol stable-carbon-isotope studies from El-Kherba, Algeria. Quaternary Science Reviews, 30: 1303–1317.
- Sahnouni, M., J. van der Made & M. Everett, 2010. Early North Africa: chronology, ecology, and hominin behavior. Insights from Ain Hanech and El-Kherba, northeastern Algeria. Quaternary International, 223-224: 436-438.
- Sahnouni, M. & J. van der Made, 2009. The oldowan in north Africa within a biochronological framework. In: K. Schick & N. Toth (eds) The cutting edge: new approeaches to the archaeology of human origins. Stone Age Institue Press - Gosport: 179-210.

Kolpochoerus from Ain Hanech compared with those of other localities (Sahnouni & Van der Made, 2009).
- Made, J. van der, 2008. Hallazgo de una tortuga gigante en Argelia. Periódico del Museo Nacional de Ciencias Naturales, 8 (octubre-diciembre 2008): 10.

Giant tortoise in the Oued Boucherit (photograph Jordi Mestre; Van der Made, 2008).
- Sahnouni, M. , D. Hadjouis, J. van der Made, A. Derradji, A. Canals , M. Medig, H. Belahrech, Z. Harichane & M. Rabhi, 2004. On the earliest human occupation in North Africa: a response to Geraads et al. Journal of Human Evolution, 46: 763-775.
- Sahnouni, M., D. Hadjiouis, J. van der Made, Abd-el-Kader Derradji, A. Canals, M. Medig, H.Belahrech, Z. Harichane, M. Raghi, 2002. Further Research at the oldowan site of Ain Hanech, Northeastern Algeria. Journal of Human Evolution, 43 (6): 925-937.
Morocco: area of Ain Beni Matar & Jerada
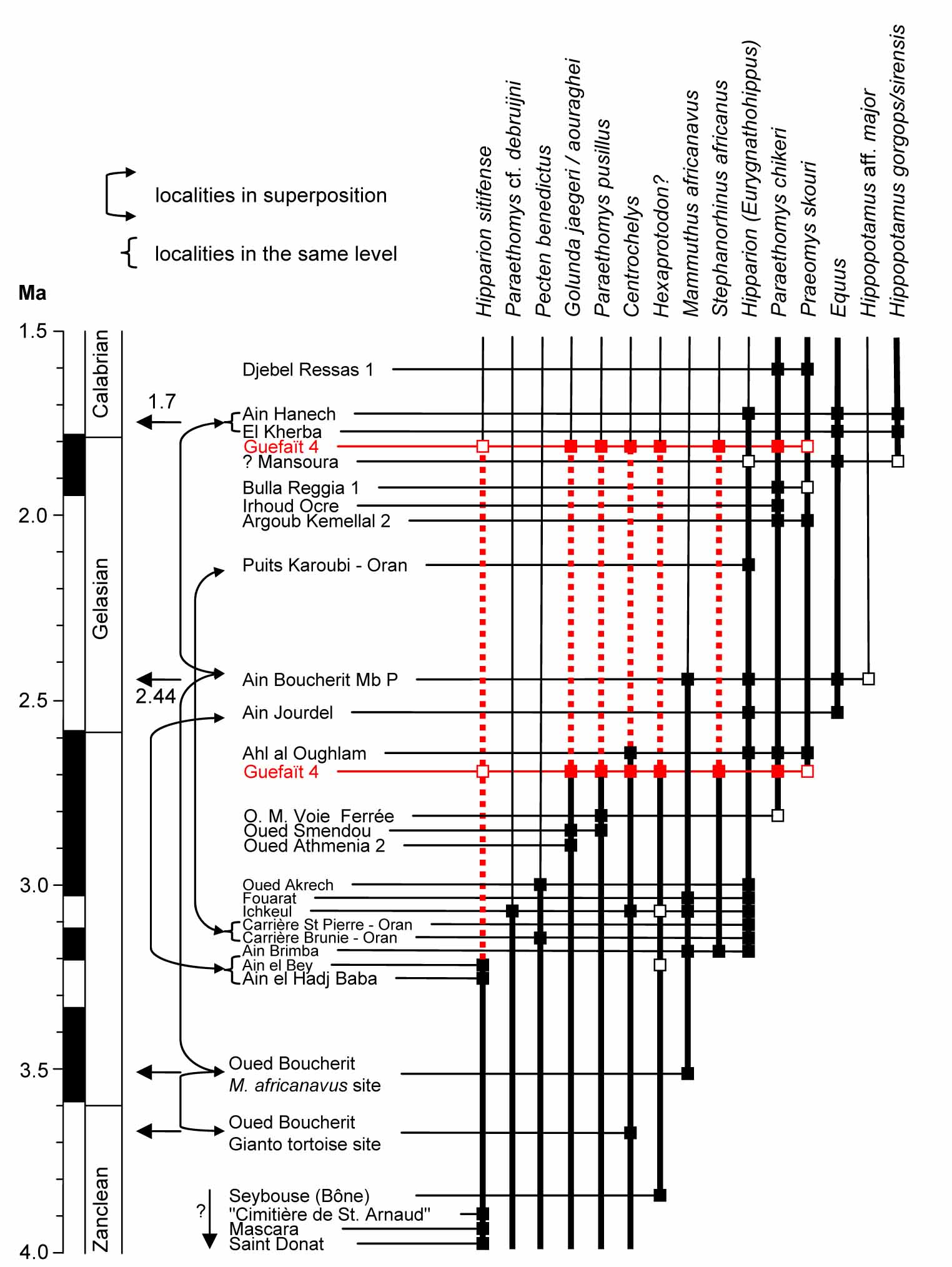
Two different hypotheses on the age of Guéfait, based on magnetostratigraphy and biostratigraphy of selected taxa (from Parés et al., 2023).
- Ramírez-Pedraza, I., C. Tornero, H. Aouraghe, F. Rivals, R. Patalano, H. Haddoumi, I. Expósito, A. Rodríguez-Hidalgo, S. Mischke, J. van der Made, P. Piñero, H.A. Blain, P. Roberts, D.K. Jha, J. Agustí, C. Sánchez-Bandera, A. Lemjidi, A. Benito-Calvo, E. Moreno-Ribas, A. Oujaa, H. Mhamdi, M. Souhir, A. Al Mahdi, M.G. Chacón & R. Sala-Ramos, 2024. Arid, mosaic environments during the Plio-Pleistocene transition and early hominin dispersals in northern Africa. Nature Communications, 15: 8393.
- Parés, J.M, H. Haddoumi, M. Duval, H. Aouraghe , C. Álvarez-Posada, S. Pla-Pueyo, A. Benito-Calvo, A. Rodríguez-Hidalgo, J. van der Made, P. Piñero, J. Agustí, A. Oujaa, R. Grün, R. Sala-Ramos, Gema Chacón, 2023. First magnetostratigraphic results in the Aïn Beni Mathar-Guefaït Basin, Northern High Plateaus (Morocco): The Pliocene-Pleistocene Dhar Iroumyane composite section. Geobios, 76: 18–37.
- Ramírez-Pedraza, I., L.M. Martínez, H. Aouraghe, F. Rivals, C. Tornero, H. Haddoumi, F. Estebaranz-Sánchez, A. Rodríguez-Hidalgo, J. van der Made, A. Oujaa, J.J. Ibáñez, H. Mhamdi, M. Souhir, A. Al Mahdi, M.G. Chacón & R. Sala-Ramos, 2023. Multiproxy approach to reconstruct fossil primate feeding behavior: Case study for macaque from the Plio-Pleistocene site Guefaït-4.2 (eastern Morocco). Frontiers in Ecology and Evolution, 11:1011208. doi: 10.3389/fevo.2023.1011208
- Sala-Ramos, R,. M. Gema Chacón, H. Aouraghe, H. Haddoumi, J.I. Morales, A. Rodríguez-Hidalgo, C. Tornero, A. Oujaa, M. Soto, M. Farkouch, A. El Mahdi, A. El Atmani, M. Duval, L. Arnold, M. Demuro, H.A. Blain, P. Piñero, F. Rivals, F. Burjachs, A. Tarriño, C. Álvarez-Posada, M. Souhir, P. Saladié, S. Pla-Pueyo, J. Cruz Larrasoaña, S. Mischke, J. Marín, E. Moreno-Ribas, A. de Lombera-Hermida, R. Bartrolí, D. Lombao, G. García-Argudo, I. Ramírez-Pedraza, C. Díez-Canseco, Sonja Tomasso, I. Expósito, E. Allué, N. Hajji, H. Mhamdi, H. Rhosne, Á. Carrancho, J.J. Villalaín, J. van der Made, A. Benito-Calvo, J. Agustí, A. Canals-Salomó & J. María Parés, 2022. Evolución del asentamiento humano en la región de Aïn Beni Mathar-Guefaït (Jerada, Marruecos oriental) - Investigaciones recientes y cartografía del poblamiento humano al norte del Sáhara. Takurunna,10-11: 179-203.
- Sala-Ramos, R., H. Aouraghe, M.G. Chacón, H. Haddoumi, J.I. Morales, A. Rodríguez-Hidalgo, C. Tornero, A. Oujaa, S. Bengamra, M. Soto, M. Farkouch, A. El Mahdi, A. El Atmani, M. Duva, L. Arnold, M. Demuro, H.A. Blain, F. Rivals, F. Burjachs, A. Tarriño, C. Álvarez-Posada , M. Souhir, P. Saladié, S. Pla-Pueyo, J. Cruz Larrasoaña, J. Marín, E. Moreno, A. De Lombera-Hermida, R. Bartrolí, D. Lombao, G. García-Argudo, I. Ramírez, C. Díez-Canseco, S. Tomasso, I. Expósito, E. Allué, H. Noureddine; H. Mhamdi, H. Rhosne, Á. Carrancho, Juan J. Villalaín, J. van der Made, Antoni Canals, A. Benito, J. Agustí & J. María Parés, 2022. Le peuplement humain pendant le Pléistocène et l'Holocène dans la Province de Jerada, Maroc Oriental. Bulletin d'Archéologie Marocaine, 27: 27-40.
- Aouraghe, H., J. van der Made, H. Haddoumi, J.i Agustí, A. Benito-Calvo, A. Rodríguez-Hidalgo, I.A. Lazagabaster, M. Souhir, H. Mhamdi, A. El Atmani, A. Ewague, R. Sala-Ramos & M.G. Chacón, 2021. New materials of the white rhinoceros Ceratotherium.simum and auerochs Bos.primigenius from a Late Pleistocene terrace of the Oued el Haï (NE Morocco) - two elements of the Maghrebi Palearctic fauna. Historical Biology,
- Alba, D.M., A. Rodríguez-Hidalgo, H. Aouraghe, J. van der Made, A. Oujaa, H. Haddoumi, P. Saladié, A. Al Mahdi, J. Marín, M. Farkouch, C. Lorenzo, S. Bengamra, E. Delson, M.G. Chacón & R. Sala-Ramos, 2021. New macaque fossil remains from Morocco. Journal of Human Evolution, 153: 102951.
Bilzingsleben & Neumark Nord (Germany)
In 1997, I joined the team that works on Bilzingsleben and Neumark Nord (Germany), led by Dietrich Mania. It was my task to study the Rhinocerotidae and Cervidae from Bilzingsleben and the Rhinocerotidae and giant deer from Neumark Nord 1. Bilzingsleben is a Middle Pleistocene site (MIS11, about 400 ka) with quite a number, but usually small, human remains and abundant fauna and lithics. Neumark Nord 1 (MIS7) was a site in a lignite mine, which is now closed. During the Pleistocene, it was small interglacial lake. Many articulated skeletons of deer, elephants, auerochsen and even of a rhinoceros are preserved. The nearby basins Neumark Nord 2 and 3 are younger.

View of the Neumark Nord lignite mine when it was still active.
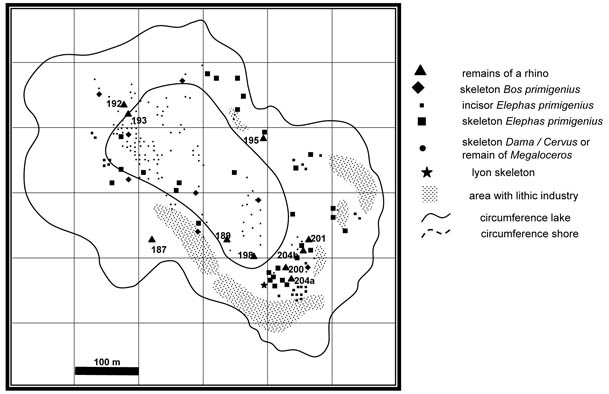
Map of the Neumark Nord 1 lake. From Van der Made (2010).

Section of Neumark Nord (from Mania, 2004, Praehistoria Thuringica, 10: 26-42). There is a controversy whether the fossils and archaeology of Neumark Nord 1 date to the Eemian (MIS5) or to the intra Saalian warm period (MIS7). In Neumark Nord 2 the Blake geomagnetic event (119-126 ka) has been detected. In the section, it can be seen that part of the sediment fill (green colour) of the Neumark Nord 1 lake ("Becken NN 1") is older than that of the Neumark Nord 2 lake ("Becken NN2") and thus older than the Blake Event and that more likely dates to MIS7. As the excavation of the mine advanced, many parallel profiles were drawn by prof. Mania, documenting the relative ages of the basin fills.
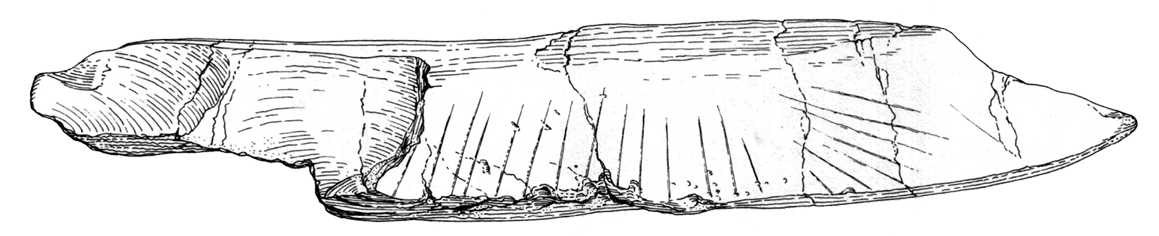
Fragment of the tibia of an elephant with regular engravings from Bilzingsleben (drawing by Mania, Praehistoria Thuringica, 1: 30-80). The pattern seems intentional.
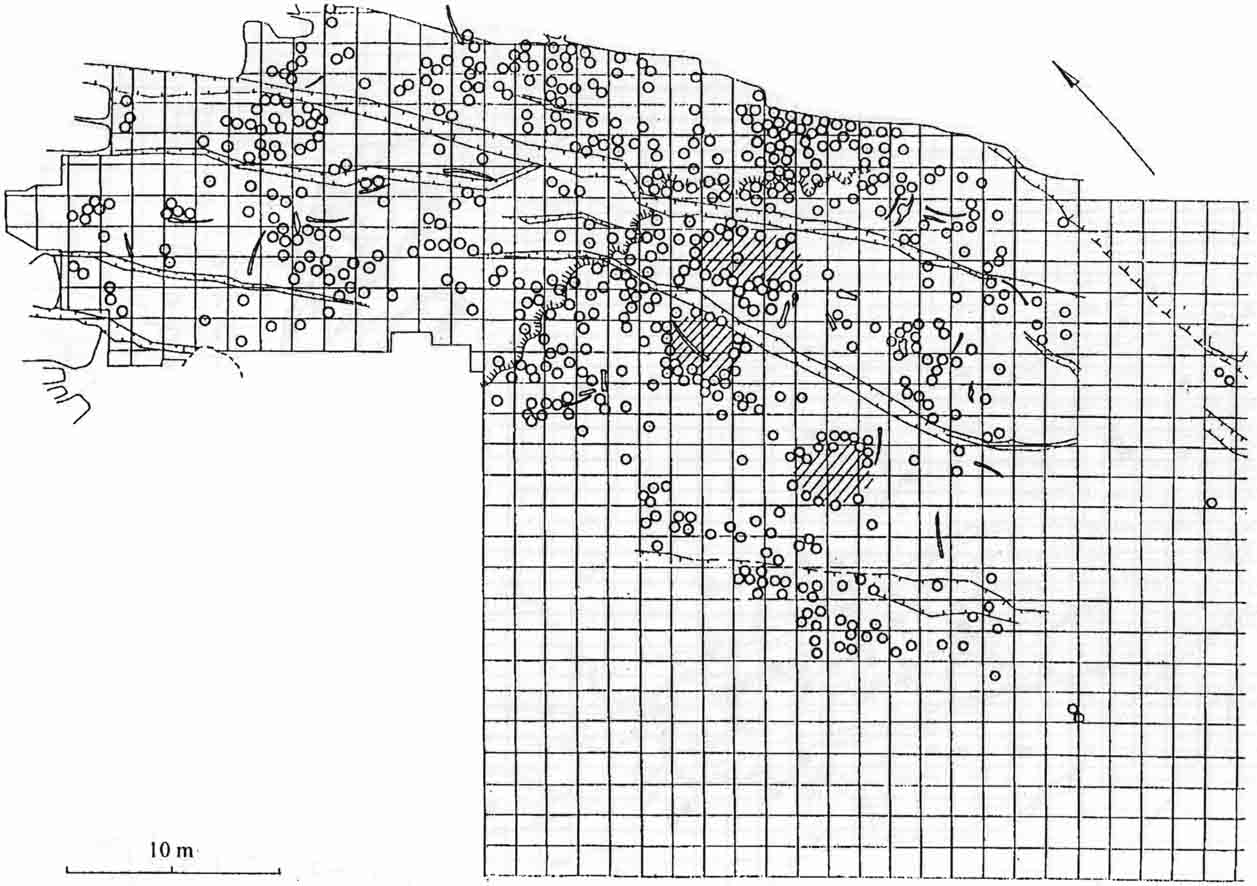
Bilzingsleben is a site in a single horizon and has been excavated in a large extension. Each square is 1.5 x 1.5 m. The dots represent the finds of large skeletal elements. (From Mania, 1997, Praehistoria Thuringica, 1: 30-80.)

Bilzingsleben: fossil bones in fine grained sediments. Though there may be areas in the site where fossils have been moved a little by water, the claim that all the site is affected by resedimentation are a bridge too far.
- Made, J. van der, 2010. The rhinos from the Middle Pleistocene of Neumark Nord (Saxony-Anhalt). Veröffentlichungen des Landesamtes für Archäologie, 62: 432-527.
- Made, J. van der, 2010. Biostratigraphy - "Large Mammals". In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 82-92.
- Made, J. van der, 2010. Giant deer. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 408-412.
- Made, J. van der & R. Grube, 2010. The rhinoceroses from Neumark-Nord and their nutrition. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 383-394.
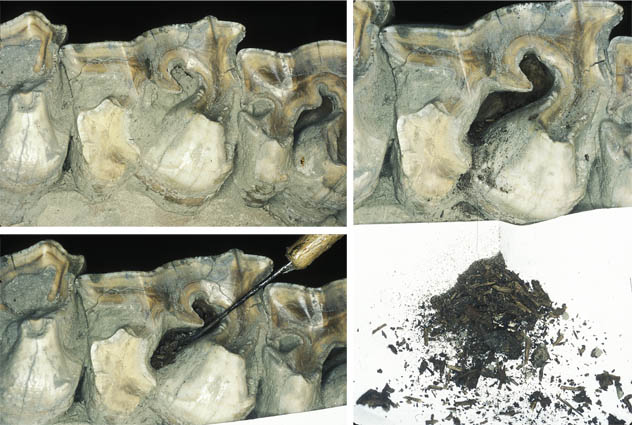
Sampling plant remains from a molar of Stephanorhinus kirchbergensis from Neumark Nord (Van der Made & Grube, 2010). These plant remains were part of the last meals of this individual.
- Made, J. van der, 2010. The evolution of the elephants and their relatives in the context of changing climate and geography. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 340-360.
- Made, J. van der, 2010. Megafauna extinctions in the wake of human dispersal. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 592-606.
- Mania, D., M. Altermann, G. Böhme, T. Böttger, E. Brühl, H.J. Döhle, K. Erd, K. Fischer, R. Fuhrmann, W.D. Heinrich, R. Grube, P.G. Karelin, J. Koller, K. V. Krementeski, T. Laurat., J. van der Made, D.H. Mai, U. Mania, R. Musil, T. Pfeiffer-Deml, E. Pietrzeniuk, T. Schüler, M. Seifert-Eulen & M. Thomae, 2010. Quartärforschung im Tagebau Neumark-Nord, Geiseltal (Sachsen-Anhalt) und ihte bisherigen Ergebnise. Veröffentlichungen des Landesamtes für Archäologie, 62: 11-69.
- Mania, D., M. Thomae, M. Alterrnann, W.D. Heinrich, J. van der Made, D.H. Mai & M. Seifert-Eulen, 2008. Zur stratigraphischen Gliederung der Saalezeit im Saalegebiet und Harzvorland. Praehistoria Thuringica, Sonderheft, 12: 1-42.
- Made, J. van der, 2003. Megaloceros giganteus from the Middle Pleistocene of Neumark Nord. Veröffentlichungen des Landesamtes für Archäologie, 57: 373-378.
- Mania, D., M. Altermann, G. Böhme, K. Erd, K. Fischer, W.D. Heinrich, C. Krementzki, J. van der Made, D.-H. Mai, R. Musil, E. Pietrzeniuk, T. Schüler, E. Vlcek & W. Steiner, 2003. Die Travertine in Thüringen und im Harzvorland. Hallesches Jahrbuch für Geowissenschaften. Reihe B: Geologie, Paläontologie, Mineralogie. Beiheft 17: 1-83.
- Mania, D., M. Algermann, G. Böhme, J. Burdukiewicz, K. Diebel, K. Erd, K. Fischer, W.D. Heinrich, E.W. Guenther, J. van der Made, D.H. Mai, U. Mania, R. Musil, T. Nötzold, E. Pietrzeniuk, W.H. Schoch, H. Süss, V. Toepfer, K. Valoch, E. Vlcek, T. Weber, G.A. Wagner, R. Mallik & T. Schüler, 2001. Homo erectus von Bilzingsleben, seine Kultur und Umwelt. In (G.A. Wagner & D. Mania, eds.) Frühe Menschen in Mitteleuropa - Chronologie, Kultur, Umwelt. Shaker Verlag, Aachen: 39-61.
- Made, J. van der, 2000. A preliminary note on the rhinos from Bilzingsleben. Praehistoria Thuringica, 4: 41-64.
- Made, J. van der, 1998. A preliminary note on the cervids from Bilzingsleben. Praehistoria Thuringica, 2: 108-122.
Iran: Kaldar Cave and other sites near Khorramabad
- Bazgir, B., E. Baharvandi, M. Azimi, J. van der Made, 2022. New chronology for the Middle Palaeolithic sequence of Kaldar Cave; Insights from the third excavation season. Journal of Archaeology and Archaeometry, 2/1: 1-11.
- Bazgir, B., A. Ollé, L. Tumung, L. Becerra-Valdivia, K. Douka, T. Higham , J. van der Made, A. Picin, P. Saladié, J.M. López-García, H.A. Blain, E. Allué, M. Fernández-García, I. Rey-Rodríguez, D. Arceredillo, F. Bahrololoumi, M. Azimi, M. Otte, E. Carbonell, 2017. Understanding the emergence of modern humans and the disappearance of Neanderthals: Insights from Kaldar Cave (Khorramabad Valley; Western Iran). Scientific Reports, 7/43460: 1-16, supplementary information: 1-16.
- Bazgir, B., M. Otte, L. Tumung, A. Ollé, S.G. Deo, P. Joglekar, J.M. López-García, A. Picin, D. Davoudi, J. van der Made, 2014. Test excavations and initial results at the Middle and UpperPaleolithic sites of Gilvaran, Kaldar, Ghamari caves and GarArjene Rockshelter, Khorramabad Valley, western Iran. Comptes Rendus Palévol, 13: 511–525.
Quibas
- Made, J. van der, J.A. Carlos Calero & M.Á. Mancheño, 2008. New material of the goat Capra alba from the Lower Pleistocene of Quibas (Spain) - notes on sexual dimorphism, stratigraphic distribution and systematics. Bolletino della Società Paleontologica Italiana, 47(1): 13-23.
- Carlos Calero, J.A., J. van der Made, M.Á.Mancheño, P. Montoya, G. Romero, 2006. Capra alba Moyà-Solà, 1987 del Pleistoceno Temprano de la Sierra de Quibas (Abanilla, Murcia). Estudios Geológicos, 62: 571-578.
- Montoya, P., M.T. Alberdi, L.J. Barbadillo, J. van der Made, J. Morales, X. Murelaga, E. Peñalver, F. Robles, A. Ruiz Bustos, A. Sánchez, B. Sanchiz, D. Soria, Z. Szyndlar, 2001. Une faune très diversifiée du Pléistocène inférieur de la Sierra de Quibas (province de Murcia, Espange). Comptes Rendus de l'Académie des Sciences, Paris, 332: 387-393.
- Montoya, P., M.T. Alberdi, A.M. Blázquez, L.J. Barbadillo, M.P. Fumaral, J. van der Made, J.M. Marín, A. Molina, J. Morales, X. Murelaga, E. Peñalver, F. Robles, A. Ruiz Bustos, A. Sánchez, B. Sanchiz, D. Soria & Z. Zsyndlar, 1999. La fauna del Pleistoceno Inferior de la Sierra de Quibas (Abanilla, Murcia). Estudios geológicos, 55(3-4): 129-164.
Madrid - Manzanares valley

Reconstruction of the Pleistocene Manzanares valley with and other fauna (by J. Gamarra; Van der Made et al., 2023).
- Martos, J.A. J. van der Made, J.J. Rodríguez-Alba, J. Gamarra, S. Rubio-Jara, J. Panera, J. Yravedra, D. Orozco, I. Claver, 2024. Vitrina CERO: Dama celiae. El gamo que habitó el valle del Manzanares (Madrid) hace 300 000 años entre cazadores achelenses. Boletín del Museo Arqueológico Nacional, 43: 321-333.
- Made, J. van der, J.J. Rodriguez Alba, J.A. Martos, J. Gamarra, S. Rubio Jara, J. Panera, J. Yravedra, 2023. The fallow deer Dama celiae sp. nov. with two.pointed antlers from the Middle Pleistocene of Madrid, a contemporary of humans with Acheulean technology. Archaeological and Anthropological Sciences, 15: 41. https://doi.org/10.1007/s12520-023-01734-3
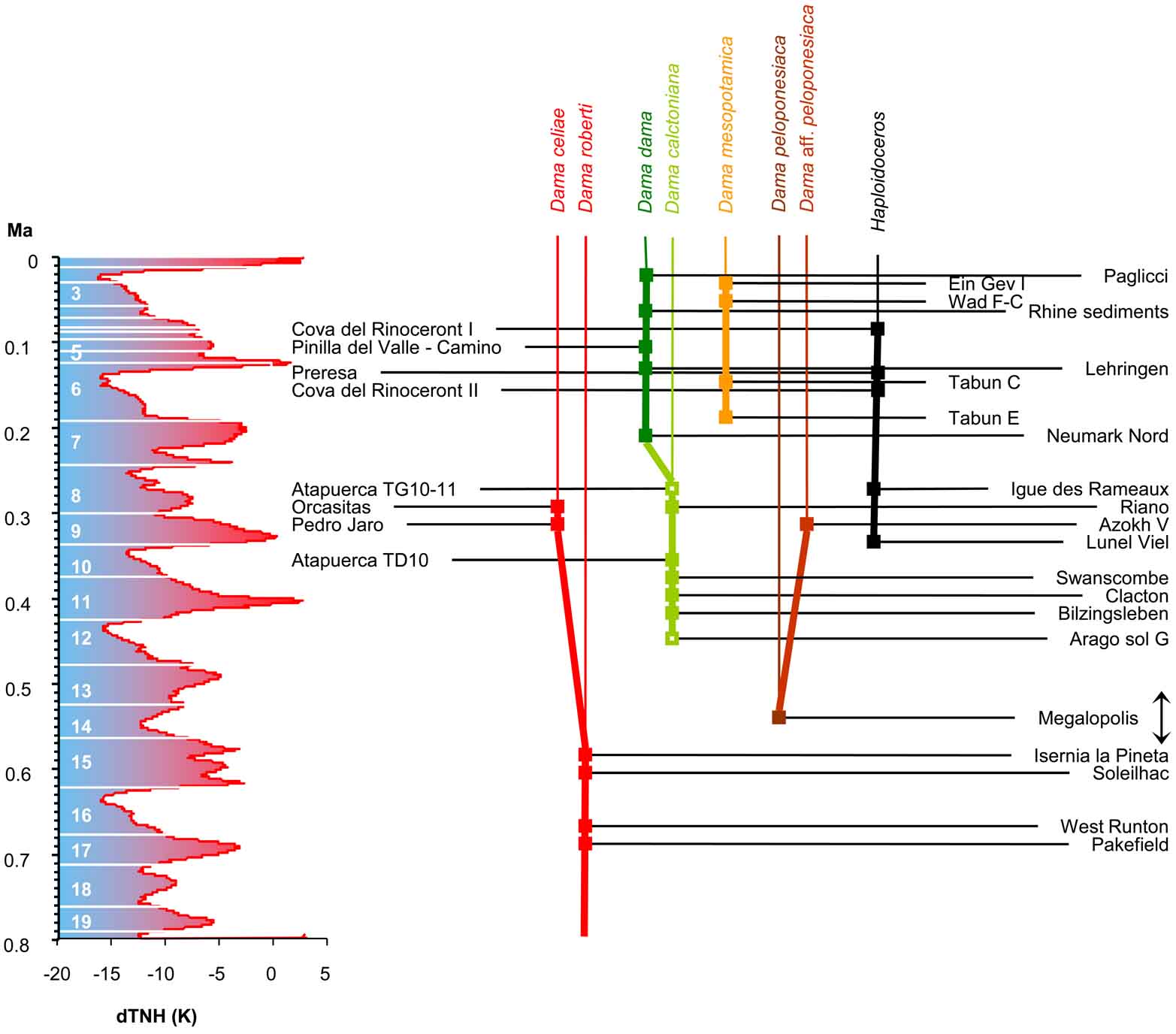
Richness of deer of the size of a fallow deer in the late Middle Pleistocene of Europe (from Van der Made et al., 2023).
- Made, J. van der, 2020. Een verdwergd reuzenhert uit de Pleistocene terrassen van de Manzanares bij Madrid. Cranium, 37/1: 41-60.
- Made, J. van der, 2019. The dwarfed "giant deer" Megaloceros matritensis n.sp. from the Middle Pleistocene of Madrid - a descendant of M. savini and contemporary to M. giganteus. Quaternary International, 520: 110-139.
- Made, J. van der, J. Panera, J. Yravedra & M.B. Ruiz Zapata, 2019. Megaloceros matritensis, un ciervo gigante que vivió en Madrid hace entre 300.000 y 400.000 años. Naturalmente, 21:40-45.
- Made, J. van der, S. Rubio Jara & J. Panera, 2019. Las Terrazas del Río Manzanares. Naturalmente, 24: 84-89.
- Yravedra, J., S. Rubio-Jara, J. Panera, J. van der Made, A. Pérez-González, 2019. Neanderthal diet in fluvial environments at the end of the Middle Pleistocene/early Late Pleistocene of PRERESA site in the Manzanares Valley (Madrid, Spain). Quaternary International, 520: 72-83.
- Made, J. van der & A.V. Mazo, 2014. Los grandes mamíferos del yacimiento de Preresa. En: I. Baquedano Beltrán & C. Laplana (eds) Haploidoceros mediterraneus una nueva especie de ciervo en el Pleistoceno Ibérico. Museo Arqueológico Regional - Alcalá de Henares: 39-53.

Homo neanderthalensis and Haploidoceros mediterraneus from the locality Preresa in the Manzanares valley. Reconstruction by Mauricio Antón.
Camp dels Ninots

Tapirus arvernensis from Camp dels Ninots. Photograph by Gerar Campeny.
- Campeny Vall-Llosera, G., B. Gómez de Soler, J. Agustí, R. Sala, O. Oms, J. van der Made, F. Burjachs, G. Jiménez, H.A. Blain, J. Claude, T. Prikryl, I. Expósito, J.J. Villalain,A. Carrancho, E. Barrón, S. Roubach, L. López-Polín, X. Bolós, G. Gómez Merino, N. Ibáñez, L. García, P. Mateos, D. Riba, R. Rosillo, A. Diaz & S. García Catalán, 2015. El Camp dels Ninots (Caldes de Malavella, la Selva): balanc de deu anys de intervencions arqueopaleontologiques. Tribuna d'Arqueologia 2012-2013: 141-163.
- Carrancho Alonso, Á, J.J. Villalaín, B. Gómez de Soler, G. Campeny Vall-llosera, M. Calvo-Rathert, O. Oms, J. van der Made & J. Agustí, 2011. Estudio magnetoestratigráfico en dos sondeos del maar del Camp dels Ninots, Caldes de Malavella (Girona, NE España). Latinmag Letters, 1(2), B20: 1-5.
- Jiménez-Moreno, G., F. Burjachs, I. Expósito, O. Oms, Á. Carrancho, J.J. Villalaín, J. Agustí, G. Campeny, B. Gómez de Soler & J. van der Made, 2013. Late Pliocene vegetation and orbital-scale climate changes from the western Mediterranean area. Global and Planetary Change, 108: 15–28.
- Gómez de Soler, B., G. Campeny Vall-Llosera, J. van der Made, O. Oms, J. Agustí, R. Sala, H.A. Blain, F. Burjachs, J. Claude, S. García Catalán, D. Riba1 & R. Rosillo, 2012. The Camp dels Ninots (NE Spain): a new key locality for the Middle Pliocene terrestrial faunas of Europe. Geologica Acta, 10(2): 1-17.
- Made, J. van der, 2010. Els macrovertebrats del Camp dels Ninots i el seu context: canvis ambientals, evolució i estructura social. En: G. Campeny Vall-llosera & B. Gómez de Soler (eds) El Camp dels Ninots - restres de l’evolució. Ayuntament de Caldes de Malavella, Caldes de Malavella, & Institut Català de Paleoecologia Humana i Evolució Social, Tarragona: 105-128.
- Made, J. van der, B. Gómez de Soler, G. Campeny Vall-Llosera, O. Oms & H.A. Blain, 2009. The large mammals of the new Pliocene fossil locality of Camp dels Ninots (Spain). Journal of Vertebrate Paleontology, 29, supplement to no. 3: 195.
- Made, J. van der, G. Campeny Vall-Llosera & B. Gómez de Soler, 2009. Camp dels Ninots, fósiles en un volcán. Periódico del Museo Nacional de Ciencias Naturales, 10 (abril-junio 2009): 4.
Vathera
Vathera, a locality on Lesbos (Greece), was a small fossiliferous lens with an extension of little more than one square metre. Fossils were recovered when a track was constructed. In 1997 the fossiliferous site was located and it was excavated from 1998 until 2000, when it was exhausted. It yielded about 550 fossils. The age is estimated to be a little more than 2 Ma. It is called the F-site to differentiate it from various other fossil occurrences in the area, which yielded fewer fossils. The recovery of good fossils of Paradolichopithecus arvernensis led in 1999 to the organisation of a workshop on Lesbos.

View of Vathera (F-site) from a distance and of the excavation, showing the excellent preservation.
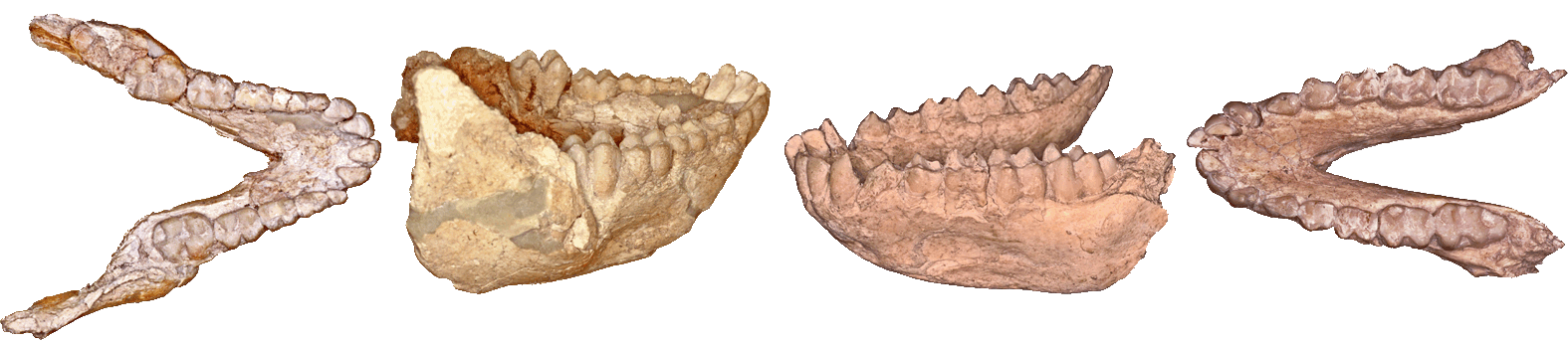
Two mandibles of Paradolichopithecus arvernensis from Vathera. The left one was my first find when starting to excavate.
- Vos, J. de, J. van der Made, A. Athanassiou, G. Lyras, P.Y. Sondaar & M.D. Dermitzakis, 2002. Preliminary note on the Plio-Pleistocene fauna from Vathera (Lesbos, Greece). Annales Géologiques des Pays Helléniques, première série, 39A: 37-70.
Others
- Rodriguez-Almagro, M, M. Arriolabengoa, M. Villalba de Alvarado, D. Arceredillo, J. Galan, M. Fernandez-Garcia, J. van der Made, J.J. Esteban, J.L. Arsuaga, J. Rios-Garaizar, N. Sala, A. Gomez-Olivencia, 2024. Palaeobiological and taphonomic analysis of a site hosting a cold-adapted fauna in Iberia: The Baio cave (Zestoa, Gipuzkoa, northern Iberian Peninsula). Geobios, 88-89: 205-217.
- Gómez-Olivencia, A.; J. Aranbarri; M. Arlegi, Lee Arnold; M. Arriolabengoa; Á. Carrancho; A. Corral; M. Demuro; M. Duval; J.J.; Esteban; M. Fernández; J. Galán; J. van der Made; V. Martínez-Pillado; C. Nuñez; A. Pantoja-Pérez; J. Rios-Garaizar; M. Rodríguez-Almagro; A. Rodríguez; N. Sala; M. Villalba de Alvarado; Antxieta Arkeologi Taldea, 2023. Cueva de Baio / Bakoko koba. Arkeoikuska, 22: 484-486.
- Villalba de Alvarado, M., E. Crégut-Bonnoure, J.L. Arsuaga, H. Collado Giraldo, J.van der Made, A. Gómez-Olivencia, 2024. Pleistocene Asian black bear (Ursus thibetanus Cuvier, 1823) in the Iberian Peninsula: new evidence and a complete review. Quaternary Science Reviews 325: 108385.
- Stefaniak, K., A. Kotowski, J. Badura, A. Sobczyk, R.K. Borówka, R. Stachowicz-Rybka, M. Moskal-del Hoyo, A. Hrynowiecka, J. Tomkowiak, J. Sawinska, B. Przybylski, D. Ciszek, W.P. Alexandrowicz, S. Skoczylas-Sniaz, U. Ratajczak-Skrzatek, A.V. Shpansky, S.A. Ilyina, P. Moska, K. Urbanski, J. Miroslaw-Grabowska, M. Niska, C. Capalbo, D. Nowakowski, J. van der Made, D. Popovic, M. Baca, E.M.E. Billia, 2023. A skeleton of peat-trapped forest rhinoceros Stephanorhinus kirchbergensis (Jäger, 1839) from Gorzów Wielkopolski, Northwestern Poland: a record of life and death of the Eemian large mammals. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 308/1: 45–77.
- Stefaniak, K., O. Kovalchuk, A. Marciszak, V. Stepanchuk, L. Rekovets, J. van der Made, V. Yanenko, A. Tsvelykh, U. Ratajczak-Skrzatek, A. Kotowski, W. Gornig, Z. Barkaszi, 2021-on line. Middle Pleistocene fauna and palaeoenvironment in the south of Eastern Europe: A case study of the Medzhybizh 1 locality (MIS 11, Ukraine). Quaternary International.
- Sobczyk,A., R.K. Borowka, J. Badura, R. Stachowicz-Rybka, J. Tomkowiak, A. Hrynowiecka, J.Slawinska, M. Tomczak, M. Pitura, M. Lamentowicz, P. Kolaczek, M. Karpinska-Kolaczek, D. Tarnawski, M. Kadej, P. Moska, M. Krapiec, K. Stachowicz, B. Bieniek, K. Siedlik, M. Bak, J. van der Made, A. Kotowski & K. Stefaniak, 2020. Geology, stratigraphy and palaeoenvironmental evolution of the Stephanorhinus kirchbergensis-bearing Quaternary palaeolake(s) of Gorzow Wielkopolski (NW Poland, Central Europe). Journal of Quaternary Science 35/4: 539-558.
- Walker, M.J., M. Haber Uriarte, A. López Jiménez, M. López Martínez, J. van der Made, M. Duval, R. Grün & I. Martín Lerma, 2020 on line. Cueva Negra del Estrecho del Río Quípar: A dated late Early Pleistocene Palaeolithic site in southeastern Spain. Journal of Paleolithic Archaeology, 3: 816–855.
- Gómez-Olivencia, A., M. Arlegi, D. Arceredillo, E. Delson, A. Sanchis, C. Núñez-Lahuerta, M. Fernández-García, M. Villalba, J. Galán, A. Pablos, A. Rodríguez-Hidalgo, M.A. López-Horgue, M. Rodríguez-Almagro, V. Martínez-Pillado, J. Rios-Garaizar & J. van der Made, 2020. The Koskobilo (Olazti, Navarre, Northern Iberian Peninsula) paleontological collection: new insights for the Middle and Late Pleistocene in Western Pyrenees. Quaternary International, 566-567: 113-140. Supplementary data.
- Gómez-Olivencia, A., M. Arlegi, D. Arceredillo, E. Delson, A. Sanchis, C. Núñez-Lahuerta, M. Fernández-García, M. Villalba, J. Galán, A. Pablos, A. Rodríguez-Hidalgo, M.A. López-Horgue, V. Martínez-Pillado, J. Rios-Garaizar, M. Rodríguez-Almagro & J. van der Made, 2019. Nuevos datos sobre el Cuaternario de Navarra: la colección paleontológica del Koskobilo (Olazti/ Olazagutía). XV Reunión Nacional del Cuaternario - Libro de resúmenes: 318-321.
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
- Walker, M.J., D. Anesin, D. Angelucci, A. Avilés-Fernández, F. Berna, A.T. Buitrago-López, J.S. Carrión, A. Eastham, Y. Fernández-Jalvo, S. Fernández-Jiménez, J. García-Torres, M. Haber-Uriarte, A. López-Jiménez, M.V. López-Martínez, I. Martín-Lerma, J. Ortega-Rodrigáñez, J.L. Polo-Camacho, S.E. Rhodes, D. Richter, T. Rodríguez-Estrella, G. Romero-Sánchez, M. San-Nicolás-del-Toro, J.L. Schwenninger, A.R. Skinner, J. van der Made, W. Zack, 2016. A View from a Cave: Cueva Negra del Estrecho del Río Quípar (Caravaca de la Cruz, Murcia, Southeastern Spain). Reflections on Fire, Technological Diversity, Environmental Exploitation, and Palaeoanthropological Approaches. Human Evolution, 31/1-2: 1-67.
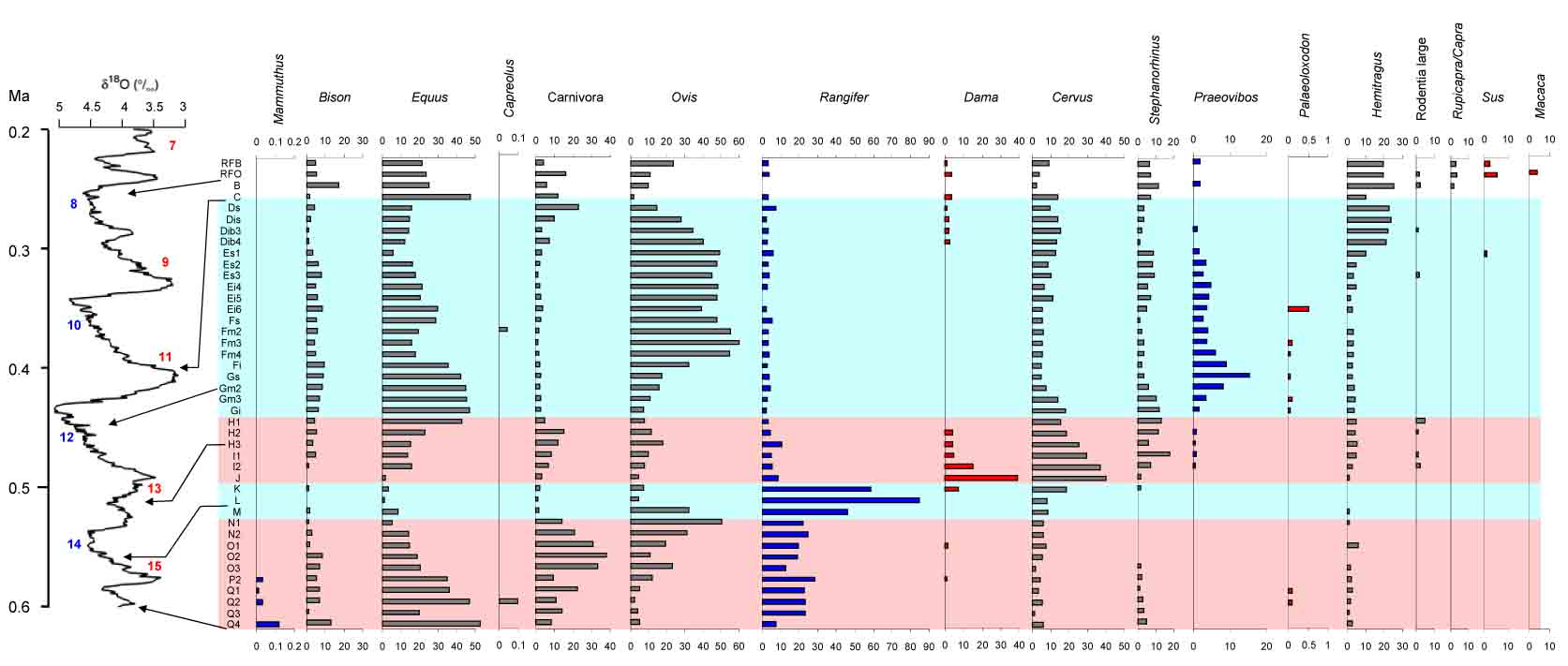
Representation of different taxa in the stratigraphic units of the Caune de l’Arago, expressed as percentages of the total of determined bones and teeth per stratigraphic unit (Van der Made, 2015).
- Made, J. van der, 2015. Caune de l'Arago: Human fossils, Archaeology and, last but not least, a long sequence of large mammals faunas. In: H. de Lumley (ed.) Caune de l'Arago, Tautavel-en-Roussion, Pyrénées-Orientales, France. Tome VI. Individualisation des unités archéostratigraphiques. CNRS Éditions, Paris: 623-628.
- Ollé, A. , P. Saladié, J. Vallverdú, I. Cáceres, J. van der Made, I. Expósito, F. Burjachs, L. López-Polín, C. Lorenzo, M. ennàsar, D. Carlos Salazar-García, C. Olària, 2014. La Cova de Dalt del Tossal de la Font. In: R. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro & J.L. Arsuaga (eds). Los cazadores recolectores del Pleistoceno y del Holoceno en Iberia y el Estrecho de Gibraltar: estado actual del conocimiento del registro arqueológico. Universidad de Burgos & Fundación Atapuerca - Burgos: 413-417.
- Ollé, A. , P. Saladié, J. Vallverdú, I. Cáceres, J. van der Made, I. Expósito, F. Burjachs, L. López-Polín, C. Lorenzo, M. Bennàsar, D. Carlos Salazar-García, C. Olària, 2014. La Cova de Dalt del Tossal de la Font. In: R. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro & J.L. Arsuaga (eds). Pleistocene and Holocene hunter-gatherers in Iberia and the Gibraltar Strait: the current archaeological record. Universidad de Burgos & Fundación Atapuerca - Burgos: 413-417.
- Gusi, F., C. Olària, A. Ollé, P. Saladié, J. Vallverdú, I. Cáceres, J. van der Made, I. Expósito, F. Burjachs, L. López-Polín, C. Lorenzo, M. Bennàsar, D.C. Salazar-García & E. Carbonell, 2013. La Cova de Dalt del Tossal de la Font (Vilafamés, Castellón): conclusiones preliminares de las intervenciones arqueológicas (1982-1987 / 2004-2012). Quaderns de Prehistòria i Arqueologia de Castelló 31: 17-38.
- López-García, J.M., G. Cuenca-Bescós, H.A. Blain, I. Cáceres, N. García, J. van der Made, J.M. Gutierrez, A. Santiago, F. Giles Pacheco, 2012. Biochronological data inferred from the Early Pleistocene Arvicolinae (Mammalia, Rodentia) of the el Chaparral site (Sierra del Chaparral, Cádiz, southwestern Spain). Journal of Vertebrate Paleontology, 32(5):1149–1156.
- Giles Pacheco, F., A. Santiago, J.M. Gutiérrez, J.M. López-García, H.-A. Blain, G. Cuenca-Bescós, J. van der Made, I. Cáceres d & N. García, 2011. The paleontological site in the Sierra del Chaparral (Villaluenga del Rosario, Cádiz-Spain). Quaternary International, 243: 92-104.
- Made, J. van der, 2011. Observations on the fauna from Terra Amata. In: H. de Lumley (ed.) Terra Amata, Nice, Alpes-Maritimes, France. Tome II. Palynologie, Anthracologie, Faunes des vertébrés, Molusques, Paléoenvironnements, Paléoanthroplogie. CNRS Éditions, Paris: 277-284.
- Fernández-Jalvo, Y., T. King, P. Andrews, L. Yepiskoposyan, N. Moloney, J. Murray, P. Domínguez-Alonso, L. Asryan, P. Ditchfield, J. van der Made, T. Torres, P. Sevilla, M. Nieto Díaz, I. Cáceres, E. Allué, M.D. Marín Monfort, & T. Sanz Martín, 2010. The Azokh cave complex: Middle Pleistocene to Holocene human occupation in the Caucasus. Journal of Human Evolution, 58(1): 103-109.
- García, N. J. Daura, M.Sanz, J. van der Made & D. Álvarez-Laó, 2010. Large Mammals from Riera dels Canyars (Gavá, Catalonia, Spain). Quaternaire, Hors série 3: 190-191.
- Saladié, P., I. Cáceres, A. Ollé, J. Vallverdú, J. van der Made, J.Mª Vergès, M.LL. Bennàsar, L. López-Polín, C. Lorenzo, I. Expósito, F. Burjachs, C. Olària, F. Gusi & E. Carbonell, 2010. Primeros resultados tafonómicos de las asociaciones fósiles de la Cova de Dalt del Tossal de la Font (Vilafamés, Castellón). Zona Arqueológica, 13: 526-537.
- Fernández-Jalvo, Y., T. Hovsepian-King, N. Moloney, L. Yepiskoposyan, P. Andrews, J. Murray, V. Safarian, L. Asryan, M. Nieto Díaz, P. Domínguez Alonso, M.ª D. Marín Monfort, E. Mkrtichyan, C. Smith, V. Bessa Correia, P. Dietchfield, E. M. Geigl, J. van der Made, T. Torres, L. Scott, E. Allue, I. Cáceres, P. Sevilla, K. Hardy, R. Grün, A. Melkonyan, R. Campos, T. Sanz Martín, H. Hayrabetyan y G. Balasanyan, 2009. Azokh Caves Excavations -2006. Middle-Upper palaeolithic transition n Nagorno-Karabagh. En: J. Lobón-Cerviá & J. Morales (eds.) Notas para la historia reciente del Museo Nacional de Ciencias Naturales. Homenaje a María Dolores Soria Mayor. Monografías del Museo Nacional de Ciencias Naturales, 24. Consejo Superior de Investigaciones Científicas, Madrid: 325-338, figuras pp. 403-410.
- Corchón, Mª.S., Ana Mateos, E. Álvarez Fernández, E- Peñalver, X. Delclòs, J. van der Made, 2008. Ressources complémentaires et mobilité dans le Magdalénien cantabrique. Nouvelles données sur les mammifères marins, les crustacés, les mollusques et les roches organogènes de la Grotte de Las Caldas (Asturies, Espagne). L’Anthropologie ,112: 284–327.
- Cuenca-Bescós, G., M.T . Alberdi, J.J. Canudo, N . García, .J .C. Gordillo, J. van der Made, D . Ramón & C. J. Rubio, 2005. Los mamíferos del Pleistoceno Medio de la Cueva de los Huesos de Obón (Parque Cultural del Río Martín, Teruel). Geogaceta, 38: 119-122.
- Sánchez, A., S. Fraile, J. van der Made, J. Morales, V. Quiralte, M.J. Salesa, I.M. Sánchez, B. Sanchiz, D. Soria, J. Jiménez, L.J. Barbadillo, C. Laplana, Z. Szyndlar, 2005. Primeros datos faunísticos del Neolítico madrileño: la cueva de la Ventana (Torrelaguna, Madrid). En P.Arias Cabal, R. Ontañón Peredo, C. García-Moncó Piñeiro (eds.) III Congreso del Neolítico en la Península Ibérica. Monografias del Instituto Internacional de Investigaciones Prehistóricas de Cantabria, 1: 155-165.
- Made, J. van der, 2004. Table 1 Taxonomic identification of fossils recovered from Azokh Cave. In: Y. Fernández Jalvo, T. King, P. Andrews, N. Moloney, P. Ditchfield, L. Yepiskoposyan, V. Safarian, M. Nieto Díaz & A. Melkonyan. Azakoh Cave and Northern Armenia. Zona Arqueologica, 4(2): 158-168.
- Canals, A., J. van der Made, I. Sauceda & E. Carbonell, 2003. El conjunto paleontológico de la Cueva de Maltravieso (Cáceres, Spain): un nuevo yacimiento del Pleistoceno. XI Reunión Nacional de Cuaternario. Oviedo, 2-4/7/2003: 313-320.
- Mazo, A.V., J. van der Made, A. Arribas & A. Sánchez, 2003. Hace 3 millones de años. Fundación de Cultura y Deporte de Castilla-La Mancha & Junta de Comunidades de Castilla-La Mancha, Ciudad Real: 1-55.
2) Evolution, systematics and biogeography of Suoidea and other groups
Suoidea
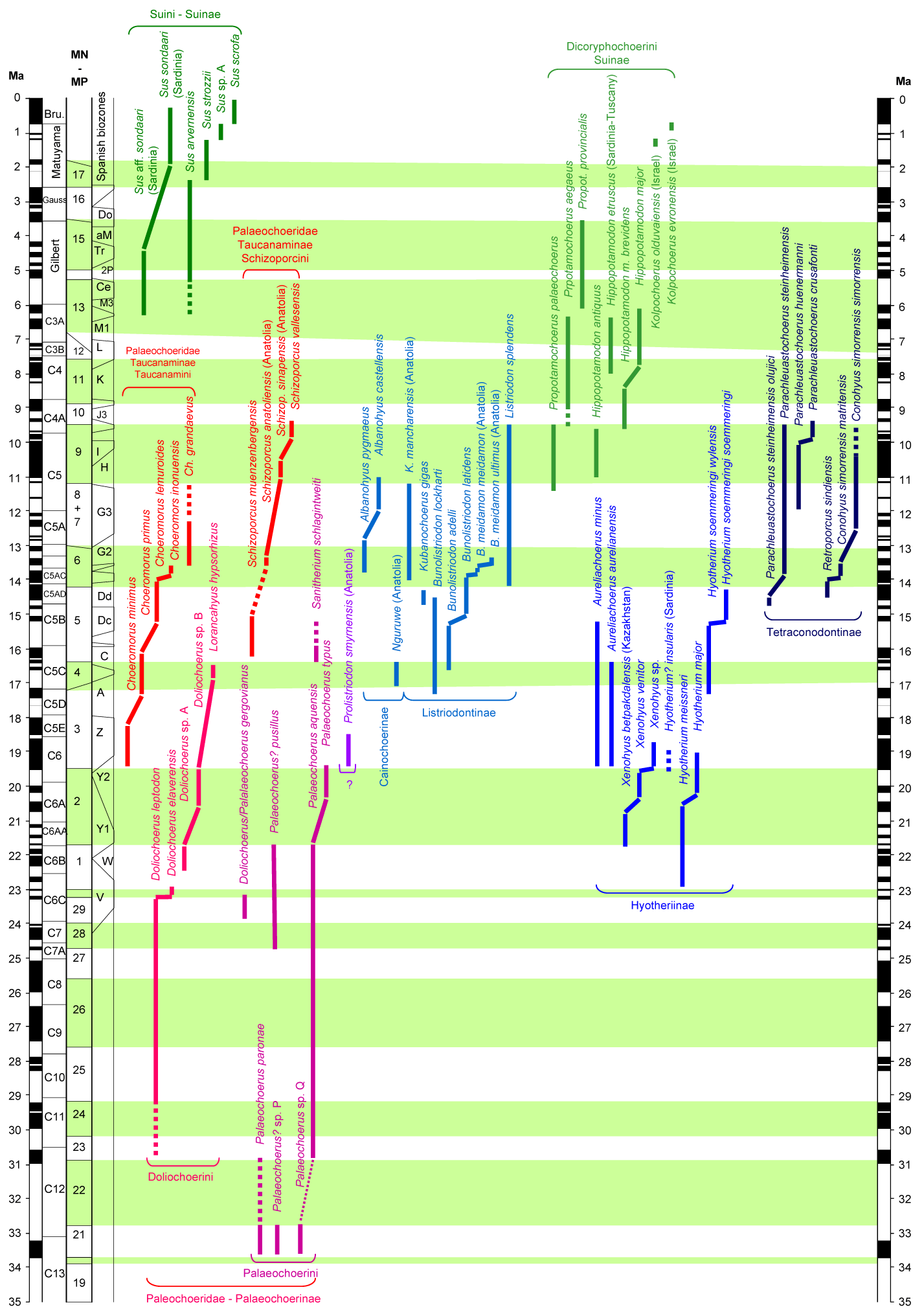
A range chart of the west Eurasian Suoidea (based on Van der Made, 2022).
- Made, J. van der, 2025. Did human dispersal into Europe cause the continent-wide extinction of the pig Sus strozzii at 1.8 Ma?—Review of a Debate. Quaternary, 8/26.
- Made, J. van der, 2023. The morphology of the upper molar roots does not provide evidence that Palaeochoerus typus belongs to the Suidae - relevance for the origin of the Suidae and the classification of the Suoidea. Revue de Paléobiologie, 42(2): 337-345.
- Made, J. van der, 2022. An updated range chart for the Oligocene to recent west Eurasian Suoidea. Variation in species richness. Paleontologia i Evolució, memòria especial 9: 34-36.
- Made, J. van der, D. Choudhary, N.P. Singh, K. M. Sharma, N.A. Singh & R. Patnaik, 2022. Listriodon dukkar sp. nov. (Suidae, Artiodactyla, Mammalia) from the Late Miocene of Pasuda (Gujarat, India): the decline and extinction of the Listriodontinae. Paläontologische Zeitschrift, 96: 355–383.
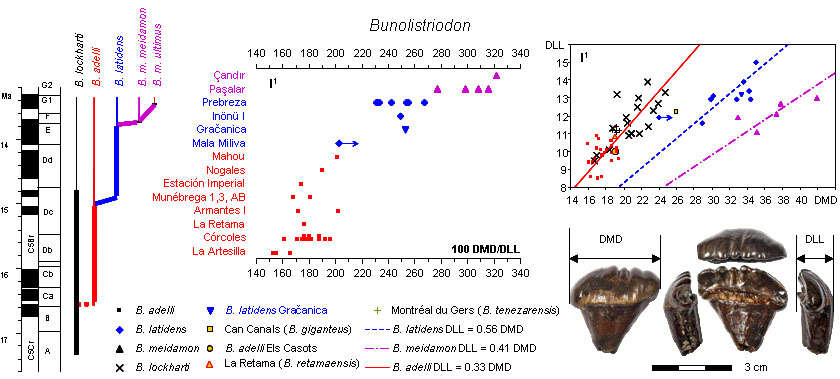
The evolution of the first upper incisor in Bunolistriodon. Adapted from Van der Made (2020).
- Made, J. van der, 2020. The Suoidea from the Middle Miocene of Graćanica (Bugojno Basin, Bosnia, and Herzegovina) — evolution, taxonomy, and biostratigraphy. Palaeobiodiversity and Palaeoenvironments, 100: 321–349. Electronic supplementary material.
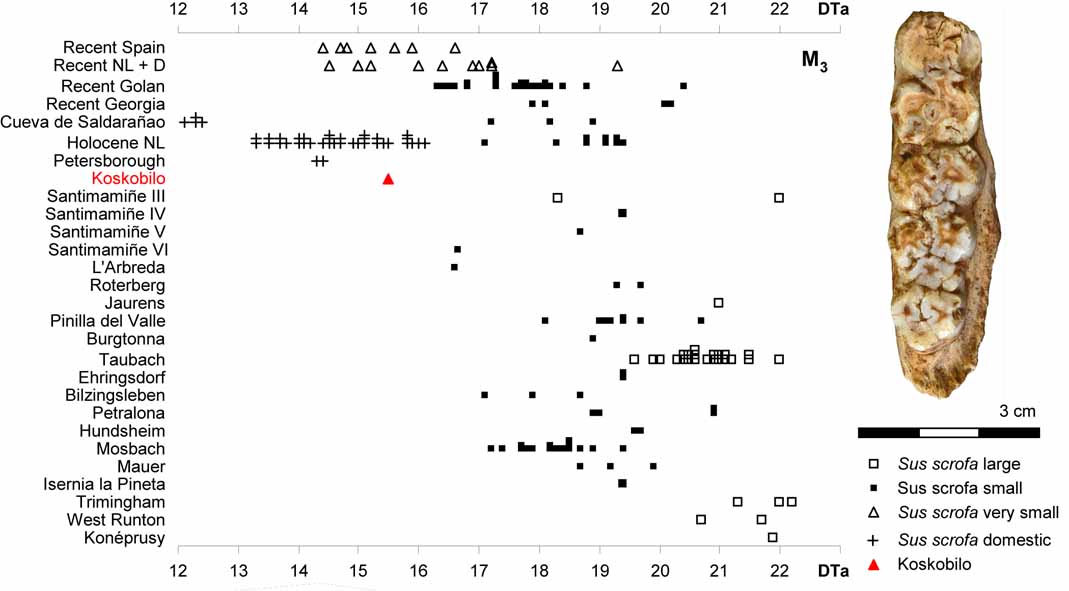
Size changes in Sus scrofa (Gómez-Olivencia et al., 2020).
- Gómez-Olivencia, A., M. Arlegi, D. Arceredillo, E. Delson, A. Sanchis, C. Núñez-Lahuerta, M. Fernández-García, M. Villalba, J. Galán, A. Pablos, A. Rodríguez-Hidalgo, M.A. López-Horgue, M. Rodríguez-Almagro, V. Martínez-Pillado, J. Rios-Garaizar & J. van der Made, 2020. The Koskobilo (Olazti, Navarre, Northern Iberian Peninsula) paleontological collection: new insights for the Middle and Late Pleistocene in Western Pyrenees. Quaternary International, 566-567: 113-140.
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
- Sahnouni, M., J.M. Parés, M. Duval, I. Cáceres, Z. Harichane, J. van der Made, A. Pérez-González, S. Abdessadok, N. Kandi, A. Derradji, M. Medig, K. Boulaghraif & S. Semaw, 2018. 1.9-million- and 2.4-million-year-old artifacts and stone tool–cutmarked bones from Ain Boucherit, Algeria. Science, 362, 1297–1301. Supplementary Materials.
- Prieto, J., P.O. Antoine, J. van der Made, G. Métais, Laq The Phuc, Quý Truong Quan, S. Schneider, Dang Ngoc Tran, Davit Vasilyan, Luong The Viet, Madelaine Böhme, 2018. Biochronological and paleobiogeographical significance of the earliest Miocene mammal fauna from Northern Vietnam. Palaeobiodiversity and Palaeoenvironments, 98/2: 287-313.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
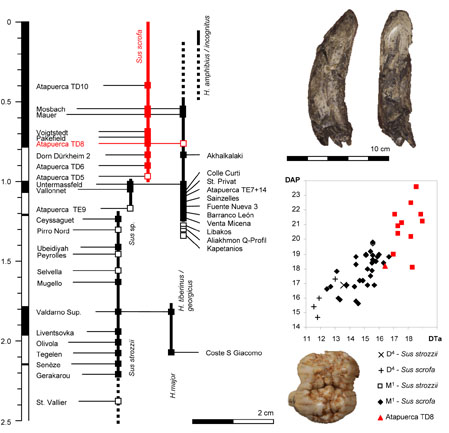
The temporal distribution of the Quaternary species of Sus and Hippopotamus in Europe (Van der Made et al., 2017).
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
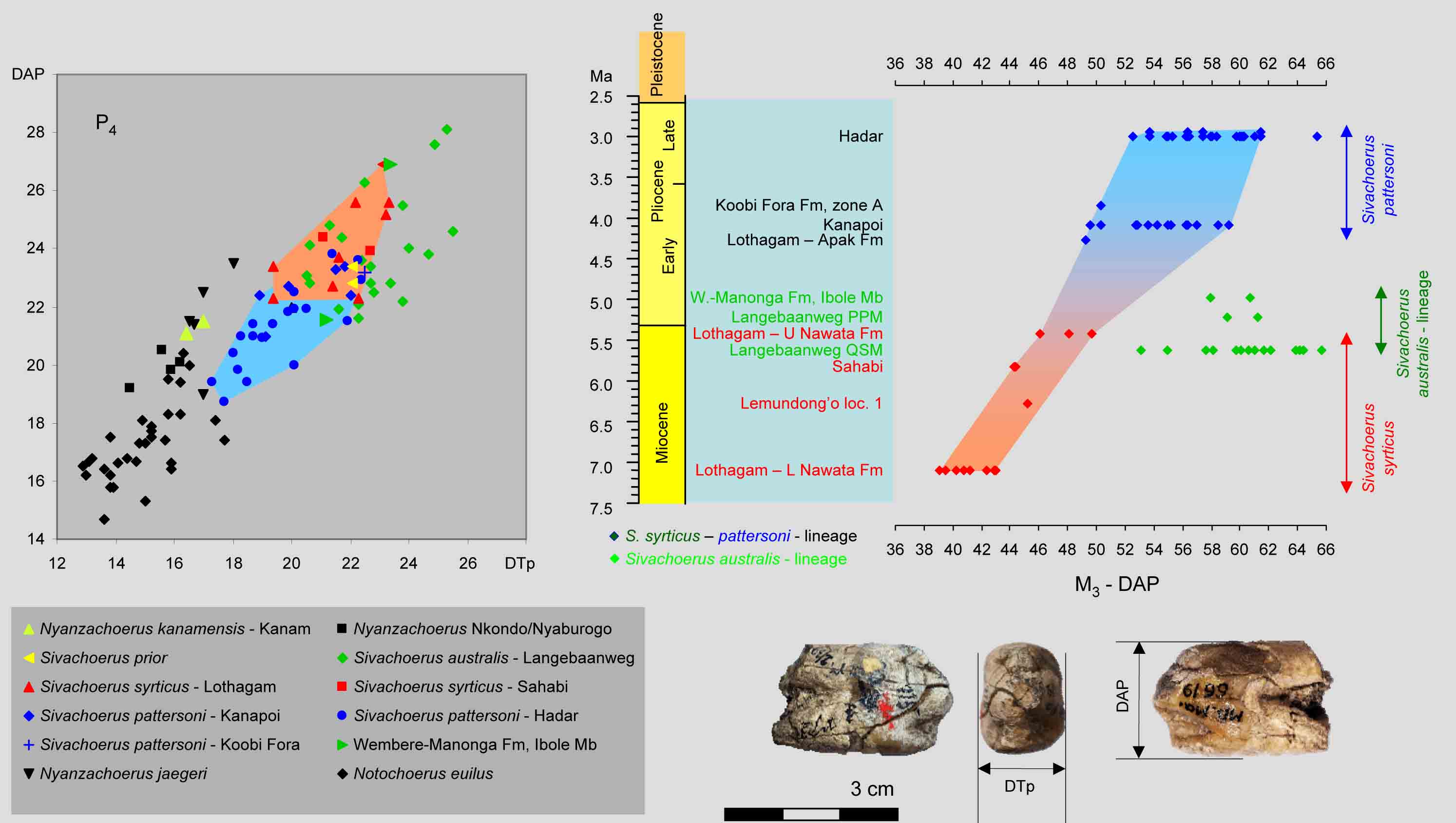
Nyanzachoerus kanamensis is a species of African pigs with narrow premolars, while Sivachoerus prior is an Indian species with wide premolars, as can be seen in the figure. It is common practice to place many African species in the genus Nyanzachoerus, however, they are morphologically more similar and related to Sivachoerus prior than to N. kanamensis. As a consequence, they should be placed in Sivachoerus. The lineage Sivachoerus syrticus-pattersoni shows size decrease of the P4 and the increase in length of the M3 with time. This lineage differs from the S. australis lineage, which at an early stage became larger with longer M3.
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364
- Made, J. van der, 2014. La evolución de los macromamíferos africanos del Plio-Pleistoceno. / The Plio-Pleistocene large mammals of Africa - Why they evolved like they did. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 137-177, English 308-323, references 360-362.
- Made, J. van der, J. Prieto, M. Aiglstorfer, M. Böhme & M. Gross, 2014. Taxonomic study of the pigs (Suidae, Mammalia) from the late Middle Miocene of Gratkorn (Austria, Styria). Palaeobiodiversity and Palaeoenvironments, 94/4: 595-617.
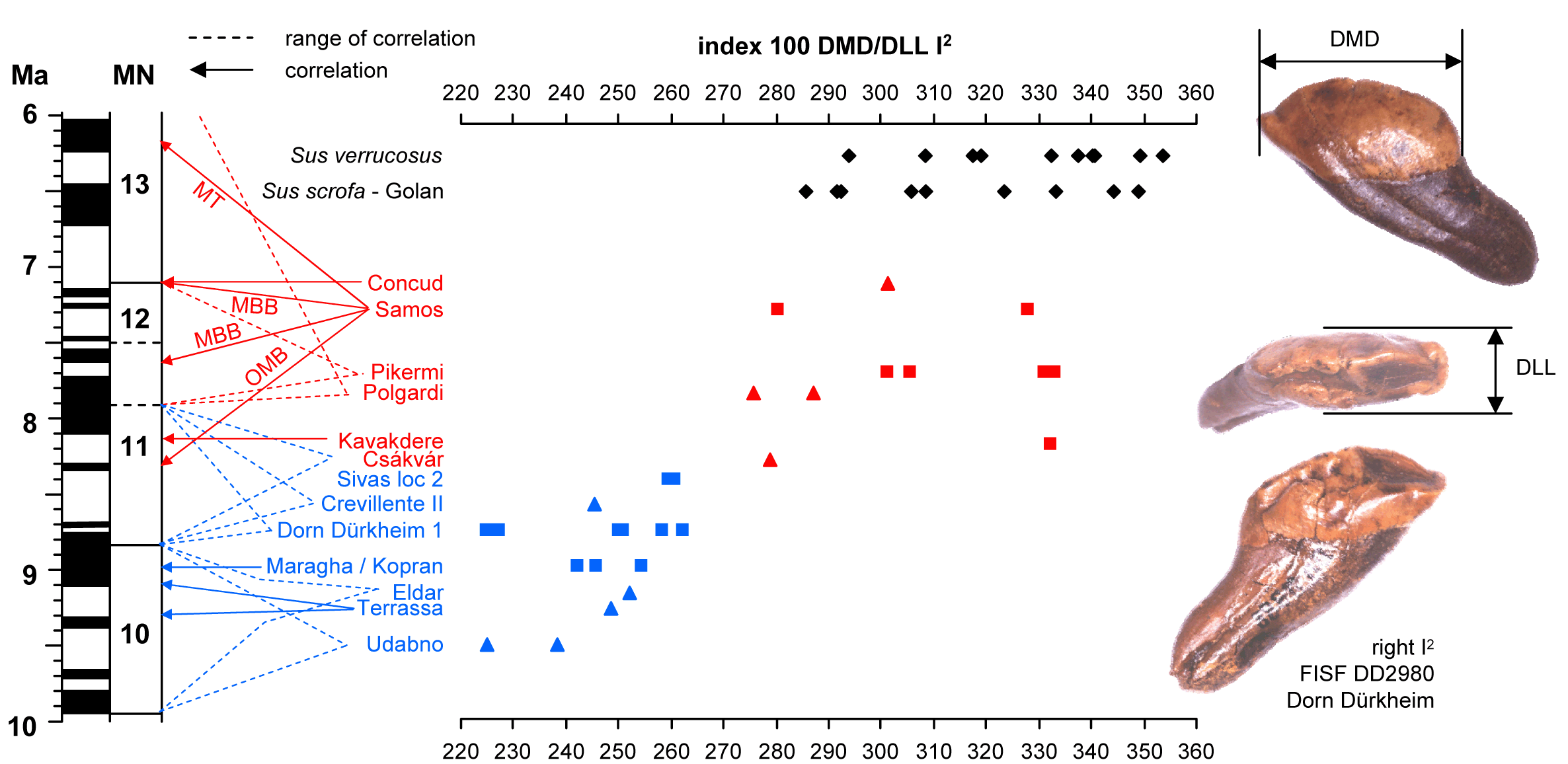
The evolutionary elongation, as indicated by the index 100DMD/DLL, of the second upper incisor of Hippopotamodon (= Microstonyx) is an adaptation to rooting and is of taxonomic and stratigraphic interest (Van der Made et al., 2013).
- Made, J. van der, E. Güleç & C.A. Erkman, 2013. Microstonyx (Suidae, Artiodactyla) from the Upper Miocene of Hayranli-Haliminhani, Turkey. Turkish Journal of Zoology, 37: 106-122.
- Made, J. van der & M. Sahnouni, 2013. Updated Plio-Pleistocene faunal lists for Ain Boucherit, Ain Hanech, and El Kherba sites, Algeria. In: M. Sahnouni (ed.) Proceedings of the International Symposium Africa, Cradle Of Humanity: Recent Discoveries. Travaux du Centre National de Recherches Préhistoriques, Anthropologiques et Historiques, Nouvelle série 18: 223-242.
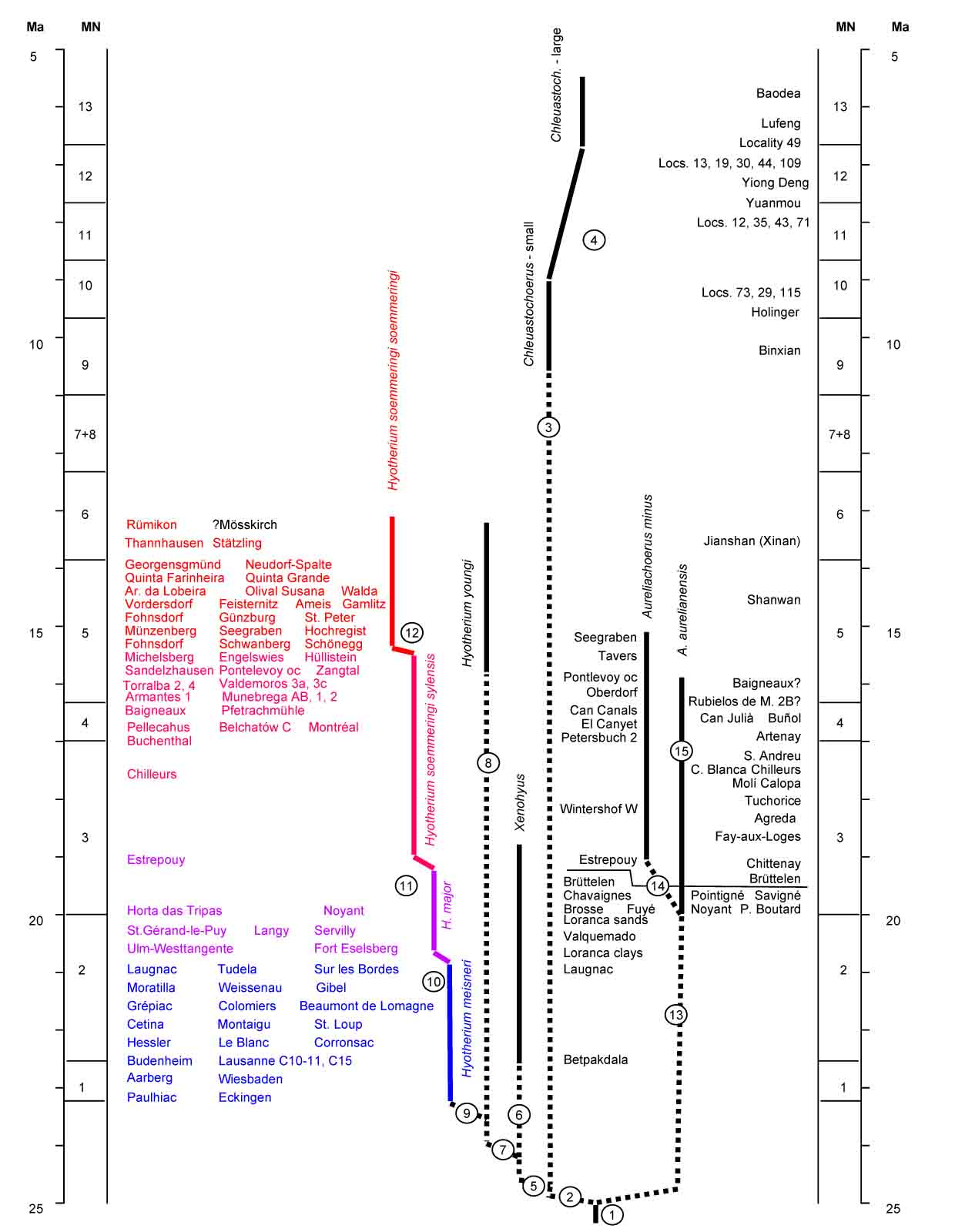
Phylogeny of the Hyotheriinae (Van der Made, 2010). The colors of the Hyotherium lineage are the same as in the following figure.
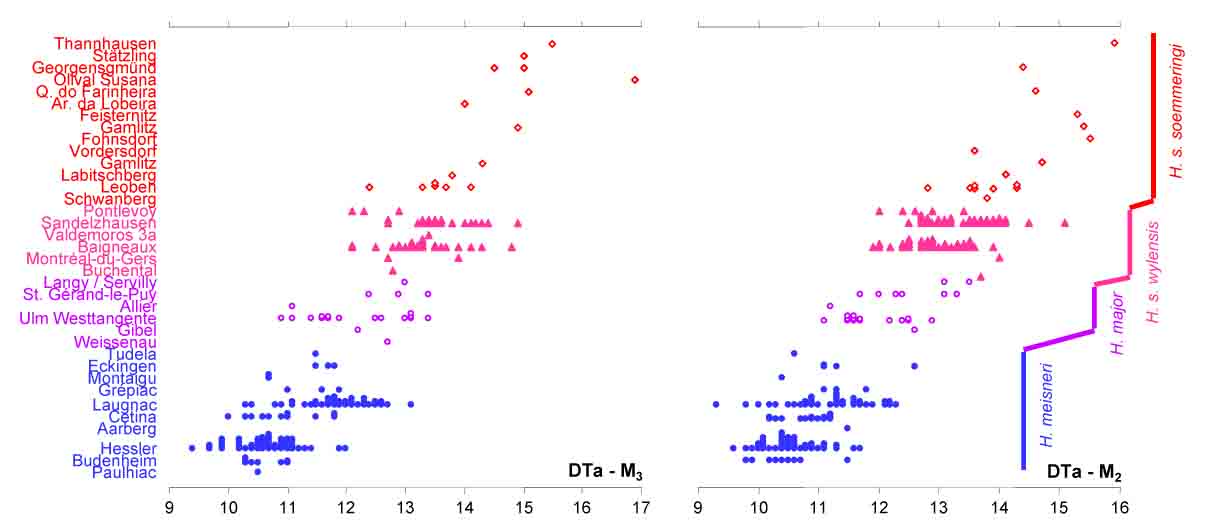
The increase in size is one of the main features in the Hyotherium lineage, here seen as an incrase in the width of the first lobes (DTa) of the first and second lower molar (Van der Made, 2010). The colors are the same as in the previous figure.
- Made, J. van der, 2010. The pigs and "Old World peccaries" (Suidae and Palaeochoeridae, Suoidea, Artiodactyla) from the Miocene of Sandelzhausen (southern Germany): phylogeny and an updated classification of the Hyotheriinae and Palaeochoeridae. Paläontologische Zeitschrift, 84: 43-121.
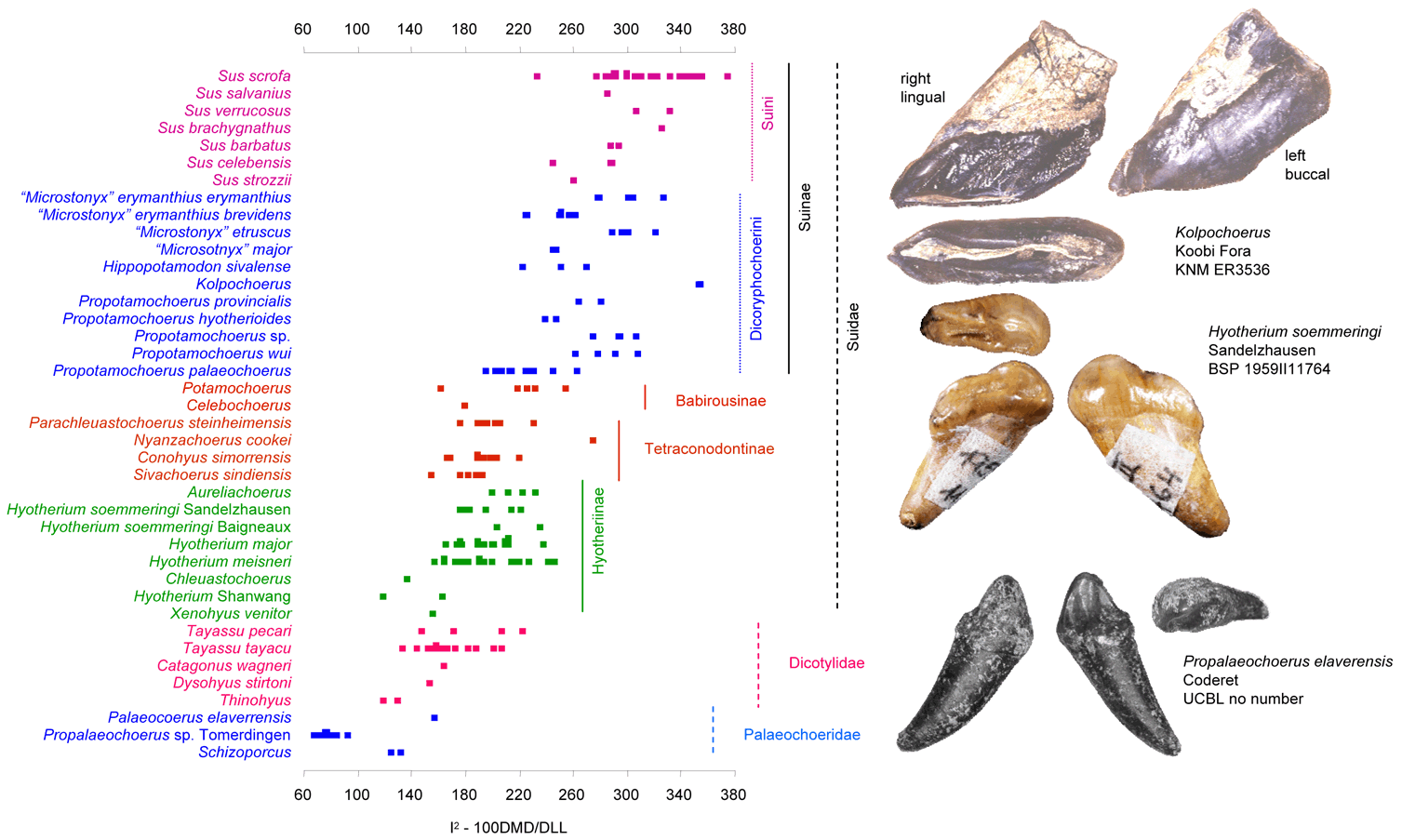
The evolutionary elongation, as indicated by the index 100 DMD/DLL, is a progressive adaptation to rooting in the Suoidea (Van der Made, 2010).
- Chit Sein, J. van der Made & G. Rössner, 2009. New material of Propotamochoerus (Suidae, Mammalia) from the Irrawaddy Formation, Myanmar. Neues Jahrbuch für Geologie und Paläontologie, 251(1): 17-31.
- Sahnouni, M. & J. van der Made, 2009. The oldowan in north Africa within a biochronological framework. In: K. Schick & N. Toth (eds) The cutting edge: new approeaches to the archaeology of human origins. Stone Age Institue Press - Gosport: 179-210.
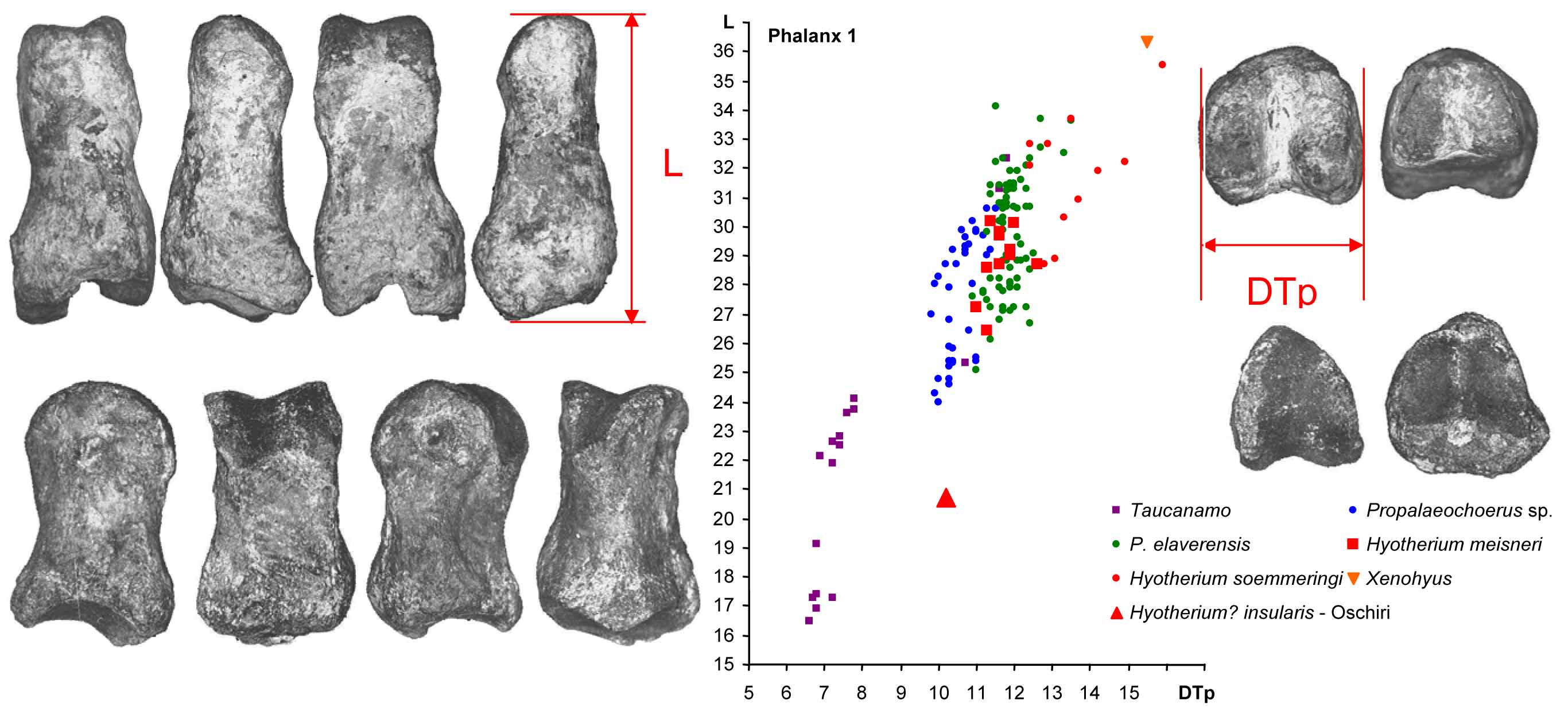
Hyotherium? insularis from Oschiri is an endemic insular species. Like many insular species, it had shortened distal limb bones, as can be seen here in tis comparison of its first phalanx to that of other species of Suoidea.
- Made, J. van der, 2008. New endemic large mammals from the Lower Miocene of Oschiri (Sardinia), observations on evolution in insular environment. Quaternary International, 182: 116-134.
- Montoya, P., L. Ginsburg, M.T. Alberdi, J. van der Made, J. Morales & M.D. Soria, 2006. Fossil large mammals from the early Pliocene locality of Alcoy (Spain) and their importance in biostratigraphy. Geodiversitas, 28(1): 137-173.
- Made, J. van der, 2005. Errata and reply to Guest Editor’s notes. Courier Forschungs-Institut Senckenberg, 254: 473-477.
- Made, J. van der, 2005. Cf. “Microstonyx” major. In F. Lacombat (ed.) Les grands Mammifères fossiles du Velay. Annales des Amis du Musée Crozatier, 13-14: 58-59.
- Sánchez, A., S. Fraile, J. van der Made, J. Morales, V. Quiralte, M.J. Salesa, I.M. Sánchez, B. Sanchiz, D. Soria, J. Jiménez, L.J. Barbadillo, C. Laplana, Z. Szyndlar, 2005. Primeros datos faunísticos del Neolítico madrileño: la cueva de la Ventana (Torrelaguna, Madrid). In P.Arias Cabal, R. Ontañón Peredo, C. García-Moncó Piñeiro (eds.) III Congreso del Neolítico en la Península Ibérica. Monografias del Instituto Internacional de Investigaciones Prehistóricas de Cantabria, 1: 155-165.
- Made, J. van der & P.E. Moullé, 2005. Listriodon splendens. In F. Lacombat (ed.) Les grands Mammifères fossiles du Velay. Annales des Amis du musée Crozetier, 13-14: 56-57.
- Sahnouni, M. , D. Hadjouis, J. van der Made, A. Derradji, A. Canals , M. Medig, H. Belahrech, Z. Harichane & M. Rabhi, 2004. On the earliest human occupation in North Africa: a response to Geraads et al. Journal of Human Evolution, 46: 763-775.
- Made, J. van der & M. Salesa, 2004. Early remains of the pig Conohyus simorrensis from the Middle Aragonian of Somosaguas near Madrid - its dispersal into Europe and evolution. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 233(2): 153-168.
- Made, J. van der, 2004. The status of Conohyus giganteus (Nyanzachoerini, Tetraconodontinae, Suidae) - Reply to Pickford (2001). Transactions of the Royal Society of Edinburgh, Earth Sciences, 94: 157-167.
- Made, J. van der & J. Morales, 2003. The pig Conohyus simorrensis from the Upper Aragonian of Alhambra, Madrid, and a review of the distribution of European Conohyus. Estudios Geológicos, 59(5-6): 303-312.
- Made, J. van der, 2003. Suoidea (pigs) from the hominoid locality of Çandir in Turkey. Courier Forschungs-Institut Senckenberg, 240: 149-179.
- Made, J. van der, 2003. Fossil Suoidea of the Miocene Sinap Formation, Turkey. In M. Fortelius, J. Kappelman, S. Sen & R.L. Bernor (eds.) Geology and paleontology of the Miocene Sinap formation, Turkey. University of Columbia Press, New York: 308-327.
- Made, J. van der, 2003. Methods in biostratigraphy - A reply to Pickford et al., 2000. Geobios, 36(2): 223-228.
- Sahnouni, M., D. Hadjiouis, J. van der Made, Abd-el-Kader Derradji, A. Canals, M. Medig, H.Belahrech, Z. Harichane, M. Raghi, 2002. Further Research at the oldowan site of Ain Hanech, Northeastern Algeria. Journal of Human Evolution, 43 (6): 925-937.
- Made, J. van der, 2001. On correct citation and on the correct interpretation of sanithere morphology. Paleontologia i Evolució, 32-33: 65-68.
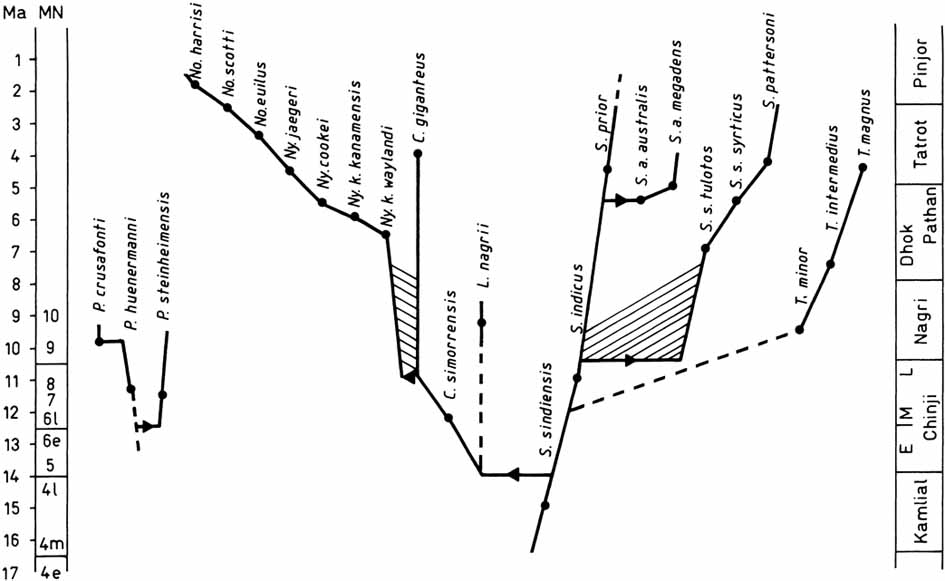
The phylogeny of the Tetraconodontinae (Van der Made, 1999).
- Made, J. van der, 1999. Biometrical trends in the Tetraconodontinae, a subfamily of pigs. Transactions of the Royal Society of Edinburgh, Earth Sciences, 89: 199-225.
- Made, J. van der, 1999. Ungulates from Atapuerca-TD6. Journal of Human Evolution, 37(3-4): 389-413.
- Made, J. van der, T. Krakhmalnaya & H. Kubiak, 1999. The pig Propotamochoerus palaeochoerus from the Upper Miocene of Grytsiv, Ukraine. Estudios geológicos, 55(5-6): 283-292.
- Made, J. van der & J. Morales, 1999. Aureliachoerus (Suidae, Mammalia) from Agreda and other Miocene localities of Spain. Geobios, 32(6): 901-914.
- Made, J. van der & F. Ribot, 1999. Additional hominoid material from the Miocene of Spain and remarks on hominoid dispersals into Europe. Contributions to Tertiary and Quaternary Geology, 36(1-4): 25-39.
- Made, J. van der & V. Tuna, 1999. A tetraconodontine pig from the Vallesian of Turkey. Transactions of the Royal Society of Edinburgh, Earth Sciences, 89: 227-230.
- Made, J. van der, 1998. Aureliachoerus from Oberdorf and other Aragonian pigs from Styria. Annalen des Naturhistorisches Museum in Wien, 99A: 225-277.
- Made, J. van der, 1998. The first described listriodont remains. Bulletin de la Société neuchâtelloise des Sciences naturelles, 121: 40-44.
- Mazo, A.V., J. van der Made, J.F. Jordá, E. Herraez, & I. Armenteros, 1998. Fauna y bioestratrigrafía del yacimiento Aragoniense de Montejo de la Vega de la Serrezuela (Segovia). Estudios Geológicos, 54(5-6): 231-248.
- Made, J. van der, M. Belinchón & P. Montoya 1998. Suoidea (Mammalia) from the Lower Miocene locality of Buñol, Valencia, Spain. Geobios, 31(1): 99-112.
- Made, J. van der, 1997. The fossil pig from the Upper Miocene of Dorn Dürkheim in Germany. Courier Forschungsinsitut Senckenberg, 197: 205-230.
- Made, J. van der 1997. Systematics and stratigraphy of the genera Taucanamo and Schizochoerus and a classification of the Palaeochoeridae (Suoidea, Mammalia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 100(1-2): 127-139.
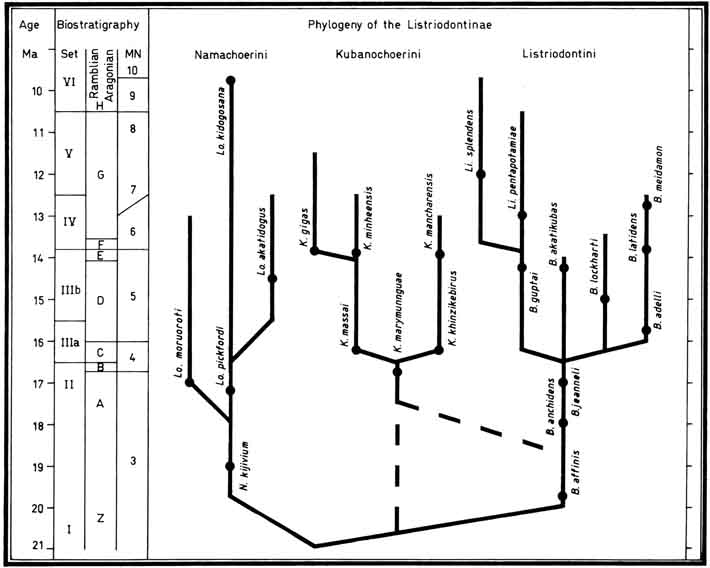
Phylogeny of the Listriodontinae (Van der Made, 1997).
- Made, J. van der 1997. On Bunolistriodon (=Eurolistriodon) and kubanochoeres. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 100(1-2): 141-160.
- Made, J. van der 1997. Los Suoidea de la Península Ibérica. In: J.P. Calvo & J. Morales (eds.) Avances en el conocimiento del Terciario Ibérico: 109-112.
- Made, J. van der, 1999. Biogeography and stratigraphy of the Mio-Pleistocene mammals of Sardinia and the description of some fossils. Deinsea, 7: 337-360.
- Fortelius, M., J. van der Made & R.L. Bernor, 1996. A new listriodont suid, Bunolistriodon meidamon sp. nov., from the Middle Miocene of Anatolia. Journal of Vertebrate Paleontology, 16(1): 149-164.
- Fortelius, M., J. van der Made & R.L. Bernor, 1996. Middle and Late Miocene Suoidea of Central Europe and the Eastern Mediterranean: Evolution, Biogeography and Paleoecology. In: R.L. Bernor, V. Fahlbusch & H.W. Mittmann (eds.) The evolution of Western Eurasian Neogene Mammal Faunas. Columbia University Press: 344-377.
- Made, J. van der, 1996. Albanohyus, a small Miocene pig. Acta Zoologica Cracoviense, 38 (1): 293-303.
- Made, J. van der, 1996. Listriodontinae (Suidae, Mammalia), their evolution, systematics and distribution in time and space. Contributions to Tertiary and Quaternary Geology, 33(1-4): 3-254. Tables 7-19 published on a microfiche.
- Made, J. van der, 1996. Pre-Pleistocene land mammals from Crete. D. S. Reese (ed.): The Pleistocene and Holocene Fauna of Crete and its First Settlers. Monographs in World Archaeology, 28: 69-79.
- Made, J. van der & K. Kowalski, 1996. Suidae (Artiodactyla, Mammalia) from the Miocene of Belchatów in Poland. Acta Zoologica Cracoviense, 38 (1): 305-310.
- Made, J. van der, 1995. When hobby horses are pigs and when opinons converge. Paleontologia i Evoluciò, 28/29: 273-275.
- Made, J. van der, 1994. Suoidea from the Lower Miocene of Cetina de Aragón, Spain. Revista Española de Paleontología, 9(1): 1-23.
- Made, J. van der & Han Defen, 1994. The Suoidea from the hominoid locality Lufeng (Yunnan, China). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 97(1): 27-82.
- Azanza, B., E. Cerdeño, L. Ginsburg, J. van der Made, J. Morales & P. Tassy, 1993. La faune des grands mammifères du Miocène Inférieur (MN 4, zone C de l'Aragonien) d'Artesilla a Villafeliche (Saragosse), Bassin de Calatayud-Teruel (Espagne). Bulletin du Muséum national d'Histoire naturelle, Paris, 4e sér. 15, section C, nos. 1-4: 105-153.
- Guan Jian & J. van der Made, 1993. The Miocene Suids from Dingjiaergou near Tongxin. Memoirs of Beijing Natural History Museum, 53: 151-199.
- Made, J. van der, P. Montoya & L. Alcalá, 1992. Microstonyx (Suidae, Mammalia) from the Upper Miocene of Spain. Geobios, 25(3): 395-413.
- Made, J. van der & S.T. Hussain, 1992. Sanitheres from the Miocene Manchar Formation of Sind, Pakistan and remarks on sanithere taxonomy and stratigraphy. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, serie B, 95(1): 81-95.
- Made, J. van der, 1991. Sexual bimodality in some recent pig populations and application of the findings to the study of fossils. Zeitschrift für Säugetierkunde, 56, 81-87.
- Made, J. van der & M. Belinchón, 1991. Korynochoerus palaeochoerus from the uppermost Miocene of Alcoy. Revista Española de Paleontología, numero extraordinario: 173-180.
- Made, J. van der, 1990. Iberian Suoidea. Paleontologia i Evolució, 23: 83-97.
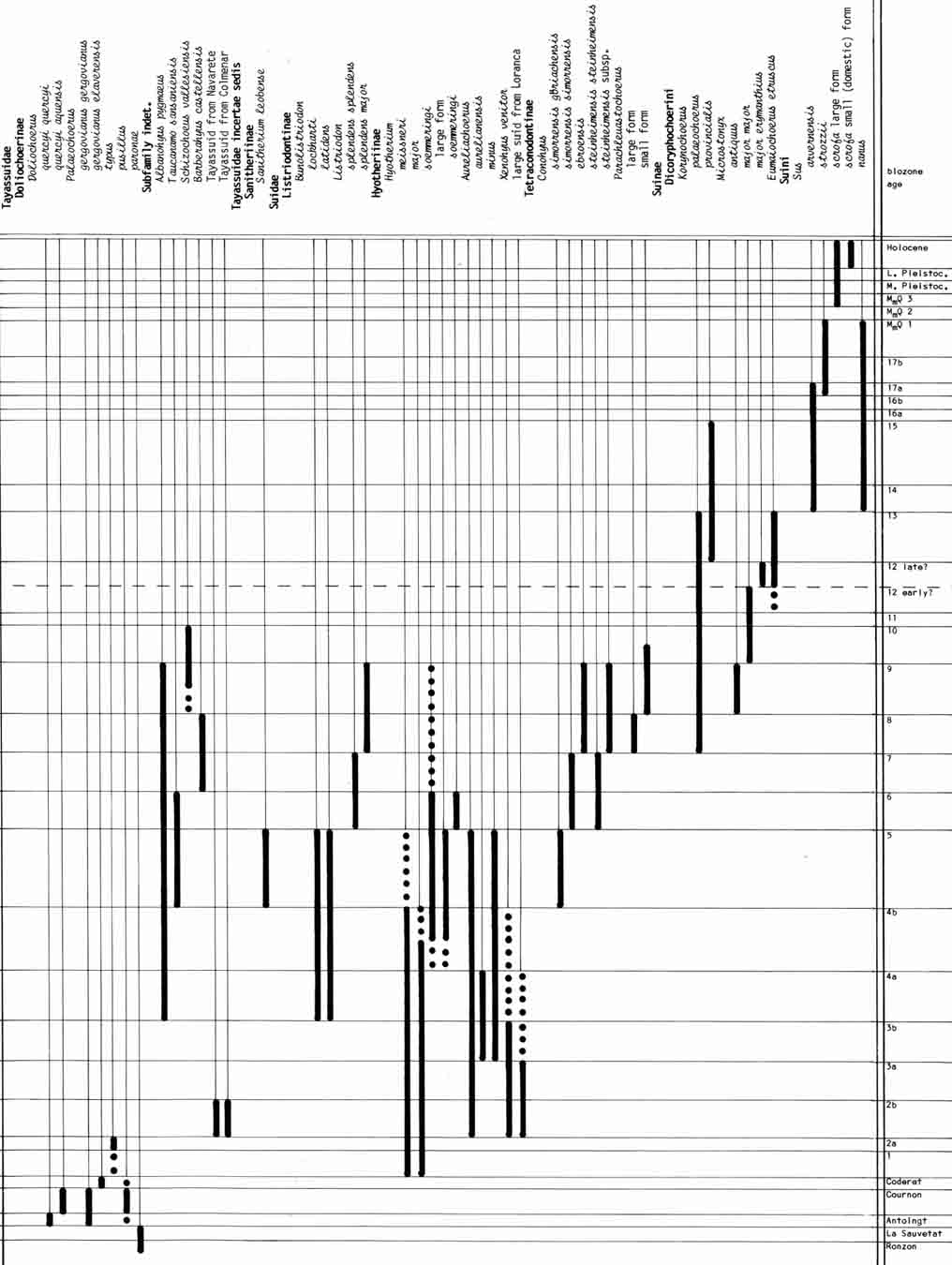
Range chart of the European Suoidea (Van der Made, 1990).
- Made, J. van der, 1990. A range chart for European Suidae and Tayassuidae. Paleontologia i Evolució, 23, 99-104.
- Made, J. van der, 1989. A Conohyus-lineage (Suidae, Artiodactyla) from the Miocene of Europe. Revista Española de Paleontología, 4: 19-28.
- Made, J. van der & S. Moyà-Solà, 1989. European Suinae (Artiodactyla) from the Late Miocene onwards. Bolletino de la Società Paleontologica Italiana, 28 (2/3): 329-339.
- Made, J. van der & S. T. Hussain, 1989. "Microstonyx" major (Suidae, Artiodactyla) from Nagri. Estudios geológicos, 45: 409-416.
- Made, J. van der, 1988. Sus nanus nov. sp. a Pliocene dwarf pig from Capo Figari (Sardinia). Bolletino della Società Paleontologica Italiana, 27 (3): 367-378, 3 pls.
Hippopotamoidea
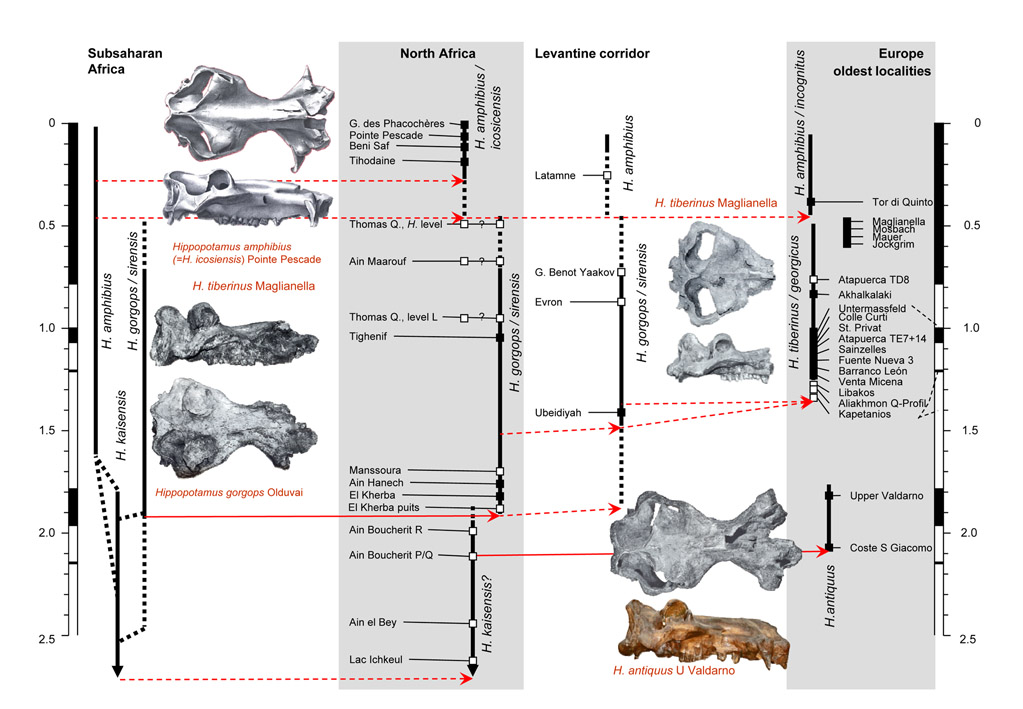
Biogeography of Hippopotamus after Van der Made et al. (2017).
- Parés, J.M, H. Haddoumi, M. Duval, H. Aouraghe , C. Álvarez-Posada, S. Pla-Pueyo, A. Benito-Calvo, A. Rodríguez-Hidalgo, J. van der Made, P. Piñero, J. Agustí, A. Oujaa, R. Grün, R. Sala-Ramos, Gema Chacón, 2023. First magnetostratigraphic results in the Aïn Beni Mathar-Guefaït Basin, Northern High Plateaus (Morocco): The Pliocene-Pleistocene Dhar Iroumyane composite section. Geobios, 76: 18–37.
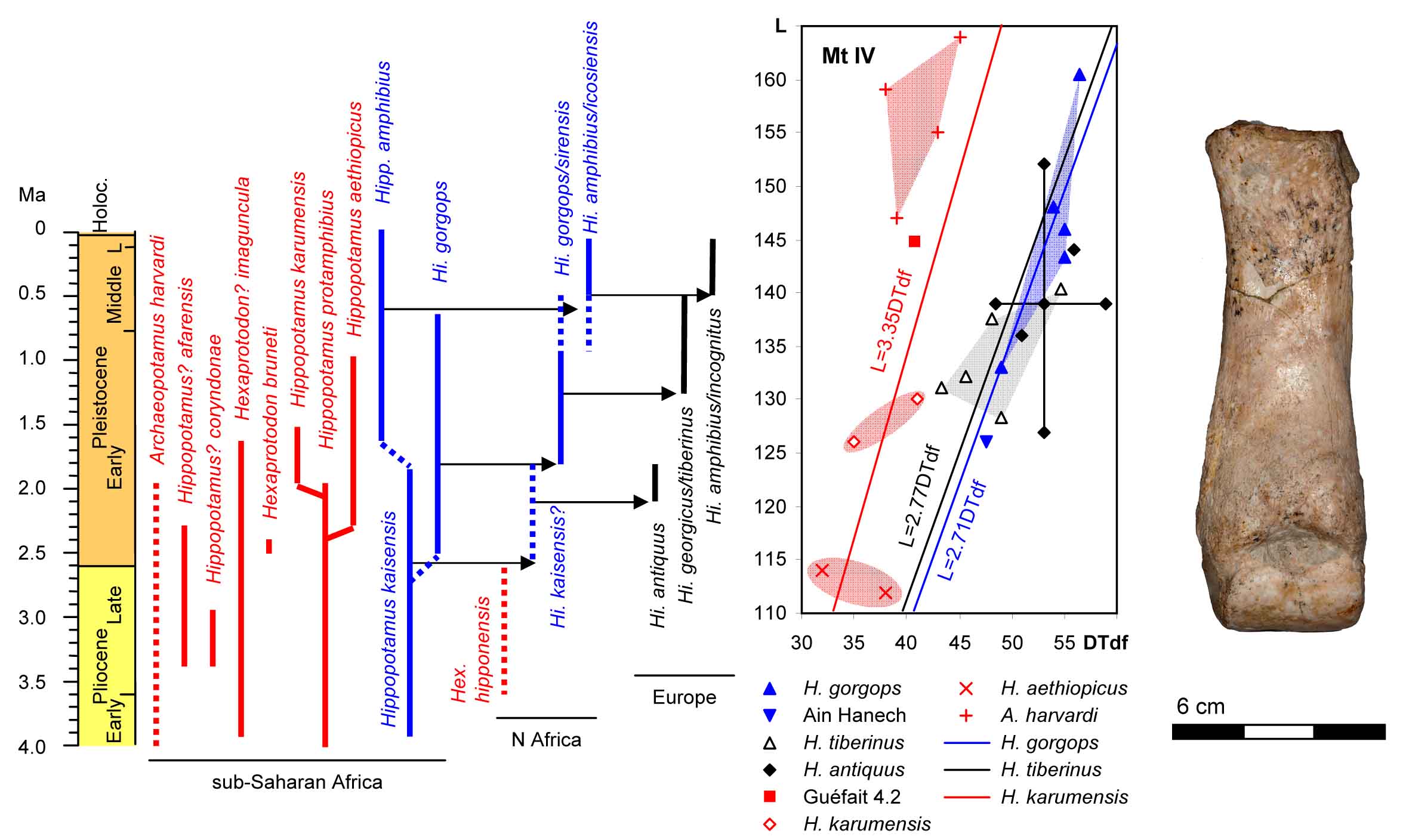
Hexaprotodon hipponensis? fourth metatarsal from Guéfait 4 (anterior view). Comparison of the length (L) and distal width (DTdf) of the fourth metatarsals of Hippopotamidae. The temporal distribution of the different species in Africa and Europe. (From Parés et al., 2023.)
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
- Made, J. van der, M. Sahnouni & Kamel Boulaghraief, 2017. Hippopotamus gorgops from El Kherba (Algeria) and the context of its biogeography. In: M. Sahnouni, S. Semaw & J. Rios Garaizar (eds.) Proceedings of the II Meeting of African Prehistory, Burgos 15-16 April, 2015. Centro Nacinonal de Investigación sobre la Evolución Humana - Burgos: 135-169.
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364.
- Ýslamoglu, Y., M. Harzhauser, M. Gross, G. Jiménez-Moreno, S. Coric, A. Kroh, F. Rögl, J. van der Made, 2010. At the Tethys - Eastern Paratethys junction: Oligocene depositional environments, paleoecology and paleobiogeography of the Thrace Basin (NW Turkey). International Journal of Earth Sciences, 99:183–200.
- Made, J. van der, 1999. Superfamily Hippopotamoidea. In (G. Rössner & K. Heissig, eds.) The Miocene land mammals of Europe. Verlag Dr. Friedrich Pfeil, München: 203-208.
Tragulidae
- Made, J. van der, 1996. Pre-Pleistocene land mammals from Crete. D. S. Reese (ed.): The Pleistocene and Holocene Fauna of Crete and its First Settlers. Monographs in World Archaeology, 28: 69-79.
Cervidae

Reconstruction of Dama celiae by J.J. Rodríguez Alba.
- Made, J. van der, J.J. Rodriguez Alba, J.A. Martos, J. Gamarra, S. Rubio Jara, J. Panera, J. Yravedra, 2023. The fallow deer Dama celiae sp. nov. with two.pointed antlers from the Middle Pleistocene of Madrid, a contemporary of humans with Acheulean technology. Archaeological and Anthropological Sciences, 15: 41. https://doi.org/10.1007/s12520-023-01734-3
- Walker, M.J., M. Haber Uriarte, A. López Jiménez, M. López Martínez, J. van der Made, M. Duval, R. Grün & I. Martín Lerma, 2020 on line. Cueva Negra del Estrecho del Río Quípar: A dated late Early Pleistocene Palaeolithic site in southeastern Spain. Journal of Paleolithic Archaeology, 3: 816–855.
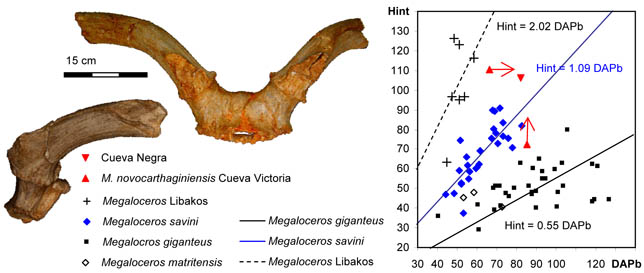
Megaloceros from Cueva Negra. After Walker et al. (2020).
- Made, J. van der, 2020. Een verdwergd reuzenhert uit de Pleistocene terrassen van de Manzanares bij Madrid. Cranium, 37/1: 41-60.
Megaloceros matritensis by Mauricio Antón.
- Made, J. van der, 2019. The dwarfed "giant deer" Megaloceros matritensis n.sp. from the Middle Pleistocene of Madrid - a descendant of M. savini and contemporary to M. giganteus. Quaternary International, 520: 110-139.
- Made, J. van der, J. Panera, J. Yravedra & M.B. Ruiz Zapata, 2019. Megaloceros matritensis, un ciervo gigante que vivió en Madrid hace entre 300.000 y 400.000 años. Naturalmente, 21:40-45.
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
- Made, J. van der & V. Dimitrijević, 2015. Eucladoceros montenegrensis n. sp. and other Cervidae from the Lower Pleistocene of Trlica (Montenegro). Quaternary International, 389: 90-118.
- Made, J. van der, 2015. The latest Early Pleistocene giant deer Megalcoeros novocarthaginiensis n. sp. and the fallow deer Dama cf. vallonnetensis from Cueva Victoria (Murcia, Spain). Mastia, 10: 269-323.
- Made, J. van der & A.V. Mazo, 2014. Los grandes mamíferos del yacimiento de Preresa. En: I. Baquedano Beltrán & C. Laplana (eds) Haploidoceros mediterraneus una nueva especie de ciervo en el Pleistoceno Ibérico. Museo Arqueológico Regional - Alcalá de Henares: 39-53.

Haploidoceros mediterraneus reconstruction by Mauricio Antón (Van der Made & Mazo, 2014).
- Made, J. van der, K. Stefaniak & A. Marciszak, 2014. The evolution and Polish fossil record of Canis, Alces, Capreolus, Megaloceros, Dama and Cervus. Quaternary International, 326-327: 406-430.
- Giles Pacheco, F., A. Santiago, J.M. Gutiérrez, J.M. López-García, H.-A. Blain, G. Cuenca-Bescós, J. van der Made, I. Cáceres d & N. García, 2011. The Early Pleistocene paleontological site in the Sierra del Chaparral (Villaluenga del Rosario, Cádiz, Southwestern Spain). Quaternary International, 243: 92-104.
- Made, J. van der, 2010. Biostratigraphy - "Large Mammals". In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 82-92.
- Made, J. van der, 2010. Giant deer. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 408-412.
- Made, J. van der & Tong H.W., 2008. Phylogeny of the giant deer with palmate brow tines Megaloceros from west and Sinomegaceros from east Eurasia. Quaternary International, 179: 135-162.
- Made, J. van der, 2006. The evolution and biogeography of the Pleistocene giant deer Megaloceros giganteus (Cervidae, Mammalia). Courier Forschungs-Institut Senckenberg, 256: 117-129.
- Made, J. van der & M.R. Palombo, 2006. Megaloceros sardus n.sp., a large deer from the Pleistocene of Sardinia. Annales Géologiques des Pays Helléniques, 41: 163-176.
- Canals, A., J. van der Made, I. Sauceda & E. Carbonell, 2003. El conjunto paleontológico de la Cueva de Maltravieso (Cáceres, Spain): un nuevo yacimiento del Pleistoceno. XI Reunión Nacional de Cuaternario. Oviedo, 2-4/7/2003: 313-320.
- Made, J. van der, 2003. Megaloceros giganteus from the Middle Pleistocene of Neumark Nord. Veröffentlichungen des Landesamtes für Archäologie, 57: 373-378.
- Made, J. van der, 2001. Les ongulés d'Atapuerca. Stratigraphie et biogéographie. L'Anthropologie, 105 (1): 95-113.
- Made, J. van der, 1999. On Euraxis, on the rules of the ICZN, on methodology. Neues Jahrbuch für Geologie und Paläontologie, Monatsheften, 1999 (11): 676-678.
- Made, J. van der, 1999. Ungulates from Atapuerca-TD6. Journal of Human Evolution, 37(3-4): 389-413.
- Made, J. van der, 1999. Artiodactylos del yacimiento mesopleistoceno de Galería (Sierra de Atapuerca). En E. Carbonell, A. Rosas & C. Diez (eds.): Ocupaciones humanas y paleoecología del yacimiento de Galería. Junta de Castilla y León: 143-167.
- Made, J. van der, 1999. Biogeography and stratigraphy of the Mio-Pleistocene mammals of Sardinia and the description of some fossils. Deinsea, 7: 337-360.
- Made, J. van der, 1998. Ungulates from Gran Dolina (Atapuerca, Burgos, Spain). Quaternaire, 9(4): 267-281.
- Made, J. van der, 1998. A preliminary note on the cervids from Bilzingsleben. Praehistoria Thuringica, 2: 108-122.
- Made, J. van der, 1996. Pre-Pleistocene land mammals from Crete. D. S. Reese (ed.): The Pleistocene and Holocene Fauna of Crete and its First Settlers. Monographs in World Archaeology, 28: 69-79.
Bovidae

One of the Alephis skeletons in the excavation at Camp dels Ninots (photograph Gerard Campeny) and reconstruction (Raúl Campuzano).
- Bartolomé, J., J. Cassinello, F. Ceacero, D. Klich, M. Komarkova, R. Taylor Lemoine, J. van der Made, F. Moran, W. Olech, J. Palau, K. Perzanowski, E. Rodríguez, 2026. Comment: Nores et al. (2024) fail in their attempt to demonstrate the inappropriateness of an eventual introduction of the European bison to Spain as a rewilding initiative. Conservation Science and Practice. 2026: 70132.
- Aouraghe, H., J. van der Made, H. Haddoumi, J.i Agustí, A. Benito-Calvo, A. Rodríguez-Hidalgo, I.A. Lazagabaster, M. Souhir, H. Mhamdi, A. El Atmani, A. Ewague, R. Sala-Ramos & M.G. Chacón, 2021. New materials of the white rhinoceros Ceratotherium.simum and auerochs Bos.primigenius from a Late Pleistocene terrace of the Oued el Haï (NE Morocco) - two elements of the Maghrebi Palearctic fauna. Historical Biology,
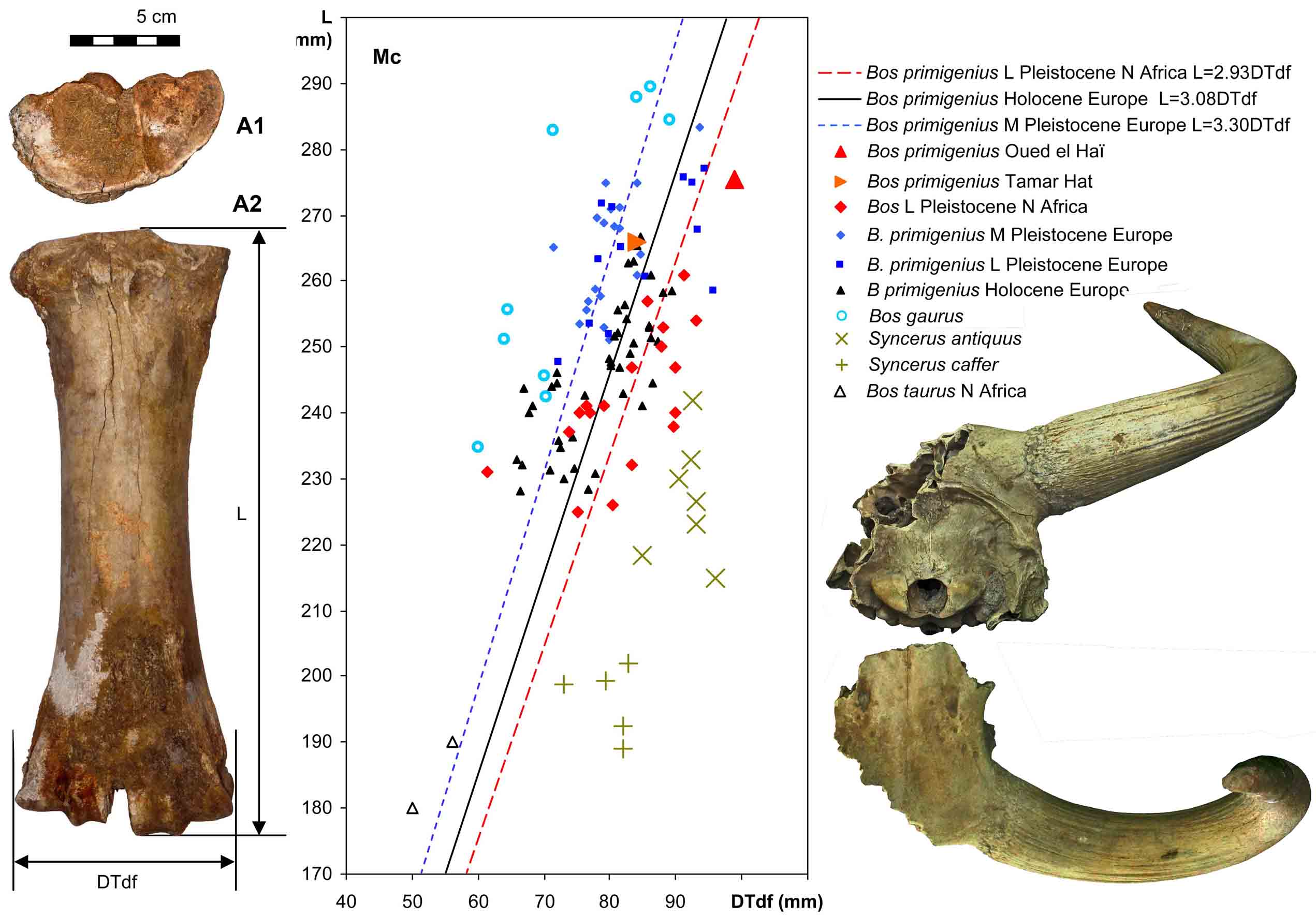
Bos primigenius evolved smaller, shorter and more robust metacarpals. This happened in Europe as well as in North Africa. Bos primigenius from Oued el Haï had already robust metacarpals, but they were still large. (From Aouraghe et al., 2021.)
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
- Azadbakht, S., A. Fazelioladi, N. Hashemi & J. van der Made, 2017. Gavkosh – a new locality yielding large mammal fossils in the Bakhtiari Formation in Lorestan Province, Iran. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 283/2: 151-159.
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
- Made, J. van der, 2014. La evolución de los macromamíferos africanos del Plio-Pleistoceno. / The Plio-Pleistocene large mammals of Africa - Why they evolved like they did. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 137-177, English 308-323, references 360-362.
- Made, J. van der, 2013. First description of the large mammals from the locality of Penal, and updated faunal lists for the Atapuerca ungulates - Equus altidens, Bison and human dispersal into Western Europe. Quaternary International, 295: 36-47.
- Made, J. van der, 2013. Morphological evidence on the origion of Bison and the Cephalophini. International conference on ruminant phylogenetics. Zitteliana, Series B, 31: 28.
- Made, J. van der & M. Sahnouni, 2013. Updated Plio-Pleistocene faunal lists for Ain Boucherit, Ain Hanech, and El Kherba sites, Algeria. In: M. Sahnouni (ed.) Proceedings of the International Symposium Africa, Cradle Of Humanity: Recent Discoveries. Travaux du Centre National de Recherches Préhistoriques, Anthropologiques et Historiques, Nouvelle série 18: 223-242.
- Gómez de Soler, B., G. Campeny Vall-Llosera, J. van der Made, O. Oms, J. Agustí, R. Sala, H.A. Blain, F. Burjachs, J. Claude, S. García Catalán, D. Riba1 & R. Rosillo, 2012. The Camp dels Ninots (NE Spain): a new key locality for the Middle Pliocene terrestrial faunas of Europe. Geologica Acta, 10(2): 1-17.
- Made, J. van der, 2012. L'Antilope Eotragus clavatus du Miocène Moyen de Sansan. Mémoires du Muséum National d'Histoire Naturelle, 203: 145-199.

Range chart of the first Bovidae (Van der Made, 2012).
- Giles Pacheco, F., A. Santiago, J.M. Gutiérrez, J.M. López-García, H.-A. Blain, G. Cuenca-Bescós, J. van der Made, I. Cáceres d & N. García, 2011. The paleontological site in the Sierra del Chaparral (Villaluenga del Rosario, Cádiz-Spain). Quaternary International, 243: 92-104.
- Made, J. van der, 2010. Els macrovertebrats del Camp dels Ninots i el seu context: canvis ambientals, evolució i estructura social. En: G. Campeny Vall-llosera & B. Gómez de Soler (eds) El Camp dels Ninots - restres de l’evolució. Ayuntament de Caldes de Malavella, Caldes de Malavella, & Institut Català de Paleoecologia Humana i Evolució Social, Tarragona: 105-128.

Alephis from Camp dels Ninots. Photograph by Gerard Campeny.
- Sahnouni, M. & J. van der Made, 2009. The oldowan in north Africa within a biochronological framework. In: K. Schick & N. Toth (eds) The cutting edge: new approeaches to the archaeology of human origins. Stone Age Institue Press - Gosport: 179-210.
- Made, J. van der, J.A. Carlos Calero & M.Á. Mancheño, 2008. New material of the goat Capra alba from the Lower Pleistocene of Quibas (Spain) - notes on sexual dimorphism, stratigraphic distribution and systematics. Bolletino della Società Paleontologica Italiana, 47(1): 13-23.
- Made, J. van der, S. Kne evic & I. Stefanovic, 2007. A mid-Miocene age for the Slanci Formation near Belgrade (Serbia), based on a record of the primitive antelope Eotragus cf. clavatus from Visnjica. Annales Géologiques de la Péninsule Balkanique, 68: 53-59.
- Carlos Calero, J.A., J. van der Made, M.Á.Mancheño, P. Montoya, G. Romero, 2006. Capra alba Moyà-Solà, 1987 del Pleistoceno Temprano de la Sierra de Quibas (Abanilla, Murcia). Estudios Geológicos, 62: 571-578.
- Made, J. van der, 2005. The fossil endemic goat Nesogoral cenisae n.sp. from Campidano Sardinia - cursorial adaptations in insular environment. Monografías Societat d’Història Natural de les Balears, 12: 347-368.
- Canals, A., J. van der Made, I. Sauceda & E. Carbonell, 2003. El conjunto paleontológico de la Cueva de Maltravieso (Cáceres, Spain): un nuevo yacimiento del Pleistoceno. XI Reunión Nacional de Cuaternario. Oviedo, 2-4/7/2003: 313-320.
- Vos, J. de, J. van der Made, A. Athanassiou, G. Lyras, P.Y. Sondaar & M.D. Dermitzakis, 2002. Preliminary note on the Plio-Pleistocene fauna from Vathera (Lesbos, Greece). Annales Géologiques des Pays Helléniques, première série, 39A: 37-70.
- Made, J. van der, 2001. Les ongulés d'Atapuerca. Stratigraphie et biogéographie. L'Anthropologie, 105 (1): 95-113.
- Montoya, P., M.T. Alberdi, A.M. Blázquez, L.J. Barbadillo, M.P. Fumaral, J. van der Made, J.M. Marín, A. Molina, J. Morales, X. Murelaga, E. Peñalver, F. Robles, A. Ruiz Bustos, A. Sánchez, B. Sanchiz, D. Soria & Z. Zsyndlar, 1999. La fauna del Pleistoceno Inferior de la Sierra de Quibas (Abanilla, Murcia). Estudios geológicos, 55(3-4): 129-164.
- Made, J. van der, 1999. Ungulates from Atapuerca-TD6. Journal of Human Evolution, 37(3-4): 389-413.
- Made, J. van der, 1999. Artiodactylos del yacimiento mesopleistoceno de Galería (Sierra de Atapuerca). En E. Carbonell, A. Rosas & C. Diez (eds.): Ocupaciones humanas y paleoecología del yacimiento de Galería. Junta de Castilla y León: 143-167.
- Made, J. van der, 1999. Biogeography and stratigraphy of the Mio-Pleistocene mammals of Sardinia and the description of some fossils. Deinsea, 7: 337-360.
- Made, J. van der & F. Ribot, 1999. Additional hominoid material from the Miocene of Spain and remarks on hominoid dispersals into Europe. Contributions to Tertiary and Quaternary Geology, 36(1-4): 25-39.
- Mazo, A.V., J. van der Made, J.F. Jordá, E. Herraez, & I. Armenteros, 1998. Fauna y bioestratrigrafía del yacimiento Aragoniense de Montejo de la Vega de la Serrezuela (Segovia). Estudios Geológicos, 54(5-6): 231-248.
- Made, J. van der, 1998. Ungulates from Gran Dolina (Atapuerca, Burgos, Spain). Quaternaire, 9(4): 267-281.
- Made, J. van der & S.T. Hussain, 1994. Horn Cores of Sivoreas (Bovidae) from the Miocene of Pakistan and Utility of their Torsion as a Taxonomic Tool. Geobios, 27(1): 103-113.
- Made, J. van der, 1989. The bovid Pseudoeotragus seegrabensis nov. gen. nov. sp. from the Aragonian (Miocene) of Seegraben near Leoben (Austria). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, serie B, 92 (3): 215-240.
Giraffidae
- Made, J. van der & J. Morales, 2011. Mitilanotherium inexpectatum (Giraffidae, Mammalia) from Huélago (Lower Pleistocene; Guadix-Baza basin, Granada, Spain) - observations on a peculiar biogegraphic pattern. Estudios geológicos, 67(2): 613-627.
- Vos, J. de, J. van der Made, A. Athanassiou, G. Lyras, P.Y. Sondaar & M.D. Dermitzakis, 2002. Preliminary note on the Plio-Pleistocene fauna from Vathera (Lesbos, Greece). Annales Géologiques des Pays Helléniques, première série, 39A: 37-70.
Camelidae

Paracamelus by Mauricio Antón.
- Made, J. van der & J. Morales, 2013. Camelids do not occur in the Late Miocene mammal locality of Çobanpinar, Turkey - omissions and contradictions. Russian Journal of Theriology, 12(1): 39-40.
- Made, J. van der, J. Morales, S. Sen & F. Aslan, 2003. A camel from Çoban Pinar. Geology and Paleontology of the Miocene Sinap Formation, Turkey. In M. Fortelius, J. Kappelman, S. Sen & R.L. Bernor (eds.) Geology and paleontology of the Miocene Sinap formation, Turkey. University of Columbia Press, New York: 328-331.
- Made, J. van der, J. Morales, S. Sen & F. Aslan, 2002. The first camel Paracamelus from the latest Miocene of Çobanpinar, Turkey. Comptes Rendus Palevol 1: 1-6.
- Made, J. van der & J. Morales, 1999. Family Camelidae. In (G. Rössner & K. Heissig, eds.) The Miocene land mammals of Europe. Verlag Dr. Friedrich Pfeil, München: 221-224.
Rhinocerotidae

The skulls of three species of rhinoceros from Neumark Nord (from left to right): Coelodonta antiquitatis, Stephanorhinus hemitoechus, and Stephanorhinus kirchbergensis (Van der Made, 2010).
- Stefaniak, K., A. Kotowski, J. Badura, A. Sobczyk, R.K. Borówka, R. Stachowicz-Rybka, M. Moskal-del Hoyo, A. Hrynowiecka, J. Tomkowiak, J. Sawinska, B. Przybylski, D. Ciszek, W.P. Alexandrowicz, S. Skoczylas-Sniaz, U. Ratajczak-Skrzatek, A.V. Shpansky, S.A. Ilyina, P. Moska, K. Urbanski, J. Miroslaw-Grabowska, M. Niska, C. Capalbo, D. Nowakowski, J. van der Made, D. Popovic, M. Baca, E.M.E. Billia, 2023. A skeleton of peat-trapped forest rhinoceros Stephanorhinus kirchbergensis (Jäger, 1839) from Gorzów Wielkopolski, Northwestern Poland: a record of life and death of the Eemian large mammals. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 308/1: 45–77.
- Aouraghe, H., J. van der Made, H. Haddoumi, J.i Agustí, A. Benito-Calvo, A. Rodríguez-Hidalgo, I.A. Lazagabaster, M. Souhir, H. Mhamdi, A. El Atmani, A. Ewague, R. Sala-Ramos & M.G. Chacón, 2021. New materials of the white rhinoceros Ceratotherium.simum and auerochs Bos.primigenius from a Late Pleistocene terrace of the Oued el Haï (NE Morocco) - two elements of the Maghrebi Palearctic fauna. Historical Biology
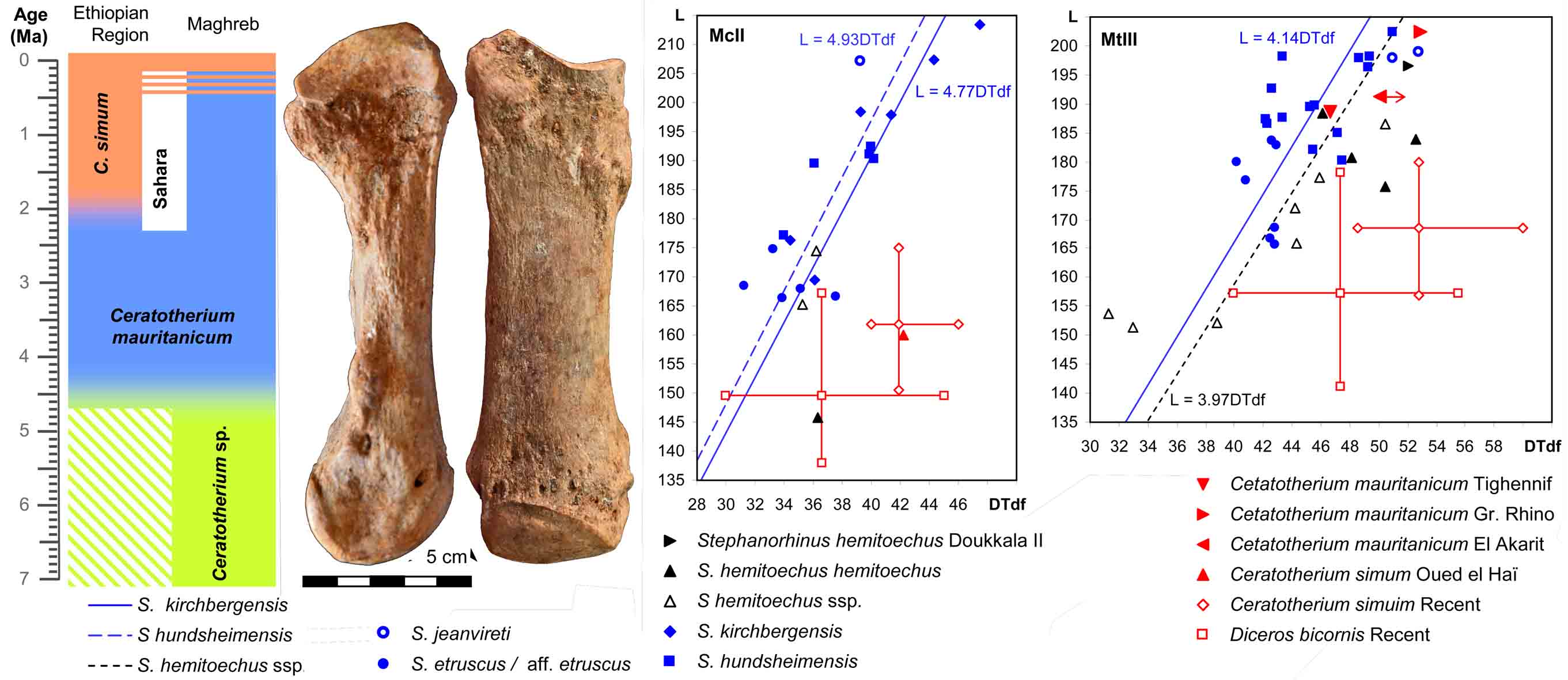
Ceratotherium simum with short and robust metapodials replaced C. mauritanicum with long and gracile metapodials in North Africa during the Late Pleistocene. Most of the many North African localities with Ceratotherium do not have material that allows a reliable identification of the species. The robust second metacarpal from Oued el Haï shows that it is one of the oldest North African C. simum. (From Aouraghe et al., 2021).
- Stefaniak, K., R. Stachowicz-Rybka, R. Krzysztof Borówka, A. Hrynowiecka, A. Sobczyk, M. Moskal-del Hoyo, A. Kotowski, D. Nowakowski, M.T. Krajcarz, E.M.E. Billia, D. Persico, E.M. Burkanova, S.V. Leschinsky, E. van Asperen, U. Ratajczak, A.V. Shpansky, M. Lempart, B. Wach, M. Niska, J. van der Made, K. Stachowicz, J. Lenarczyk & O. Kovalchuk, 2020 on line. Browsers, grazers or mix-feeders? Study of the diet of extinct Pleistocene Eurasian forest rhinoceros Stephanorhinus kirchbergensis (Jäger, 1839) and woolly rhinoceros Coelodonta antiquitatis (Blumenbach, 1799). Quaternary International, 605-606: 192-212.
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
- Piñero, P., J. Agustí, O. Oms, I. Fierro, P. Montoya, S. Mansino, F. Ruiz-Sánchez, D.M. Alba, M.T. Alberdi, H.-A. Blain, C. Laplana, J. van der Made, A.V. Mazo, J. Morales, X. Murelaga, A. Pérez-García, F. Pérez-Valera, J.A. Pérez-Valera, P. Sevilla, J.M. Soria, G. Romero, 2017. Early Pliocene continental vertebrate fauna at Puerto de la Cadena (SE Spain) and its relevelance to the marine-continental correlation of the Late Neogene of Eastern Betics. Palaeogeography, Palaeoclimatology, Palaeoecology, 479: 102–114.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
- Kirilova, I.V., O.F. Chernova, J. van der Made, V.V. Kukarskih, B. Shapiro, J. van der Plicht, F.K. Shidlovskiya, P.D. Heintzman, T. van Kolfschoten, O.G. Zanina, 2017. On the discovery of the skull of Stephanorhinus kirchbergensis (Jäger, 1839) above the Arctic Circle. Quaternary Research, 88/3: 537-550.
- Kirilova, I.V., O.F. Chernova, J. van der Made, V.V. Kukarskih, B. Shapiro, J. van der Plicht, F.K. Shidlovskiya, P.D. Heintzman, T. van Kolfschoten, O.G. Zanina, 2017. Discovery of the skull of Stephanorhinus kirchbergensis (Jäger, 1839) above the Arctic Circle – CORRIGENDUM. Quaternary Research, 88: 570.
- Made, J. van der, 2015. The rhinoceros Stephanorhinus aff. etruscus from the latest Early Pleistocene of Cueva Victoria (Murcia, Spain). Mastia, 10: 359-383.
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364.
- Gómez de Soler, B., G. Campeny Vall-Llosera, J. van der Made, O. Oms, J. Agustí, R. Sala, H.A. Blain, F. Burjachs, J. Claude, S. García Catalán, D. Riba & R. Rosillo, 2012. The Camp dels Ninots (NE Spain): a new key locality for the Middle Pliocene terrestrial faunas of Europe. Geologica Acta, 10(2): 1-17.
- Giles Pacheco, F., A. Santiago, J.M. Gutiérrez, J.M. López-García, H.-A. Blain, G. Cuenca-Bescós, J. van der Made, I. Cáceres d & N. García, 2011. The paleontological site in the Sierra del Chaparral (Villaluenga del Rosario, Cádiz-Spain). Quaternary International, 243: 92-104.
- Made, J. van der, 2010. The rhinos from the Middle Pleistocene of Neumark Nord (Saxony-Anhalt). Veröffentlichungen des Landesamtes für Archäologie, 62: 432-527.
- Made, J. van der, 2010. Els macrovertebrats del Camp dels Ninots i el seu context: canvis ambientals, evolució i estructura social. En: G. Campeny Vall-llosera & B. Gómez de Soler (eds) El Camp dels Ninots - restres de l’evolució. Ayuntament de Caldes de Malavella, Caldes de Malavella, & Institut Català de Paleoecologia Humana i Evolució Social, Tarragona: 105-128.

The temporal distribution of the western European species of Rhinocerotidae (Van der Made & Grube, 2010).
- Made, J. van der & R. Grube, 2010. The rhinoceroses from Neumark-Nord and their nutrition. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 383-394.
- Made, J. van der & P. Montoya, 2008. Rinocerontes del Pleistoceno de El Baradello, Plaça de la República y El Molinar en Alcoy, España. Recerques del Museu d’Alcoi, 16 (2007): 7-18.
- Sánchez, A., S. Fraile, J. van der Made, J. Morales, V. Quiralte, M.J. Salesa, I.M. Sánchez, B. Sanchiz, D. Soria, J. Jiménez, L.J. Barbadillo, C. Laplana, Z. Szyndlar, 2005. Primeros datos faunísticos del Neolítico madrileño: la cueva de la Ventana (Torrelaguna, Madrid). En P.Arias Cabal, R. Ontañón Peredo, C. García-Moncó Piñeiro (eds.) III Congreso del Neolítico en la Península Ibérica. Monografias del Instituto Internacional de Investigaciones Prehistóricas de Cantabria, 1: 155-165.
- Made, J. van der, 2001. Les ongulés d'Atapuerca. Stratigraphie et biogéographie. L'Anthropologie, 105 (1): 95-113.
- Made, J. van der, 2000. A preliminary note on the rhinos from Bilzingsleben. Praehistoria Thuringica, 4: 41-64.
- Made, J. van der, 1999. Ungulates from Atapuerca-TD6. Journal of Human Evolution, 37(3-4): 389-413.
- Made, J. van der, 1998. Ungulates from Gran Dolina (Atapuerca, Burgos, Spain). Quaternaire, 9(4): 267-281.
- Made, J. van der, 1998. Vertebrates from the open-cast mine Oberdorf (Western Styria Basin, Austria). Mammalia 8: Rhinocerotidae. Annalen des Naturhistorisches Museum in Wien, 99A: 163-168.
Equidae

Reconstruction of Equus altidens in the landscape of Atapuerca TD6 by Mauricio Antón (Bermúdez de Castro et al., 1999).
- Made, J. van der Made, K. Boulaghraief, R. Chelli- Cheheb, I. Cáceres, Z. Harichane & M. Sahnouni, 2022. The last North African hipparions – hipparion decline and extinction follows a common pattern. Neues Jahrbuch fürGeologie und.Paläontologie, Abhandlungen, 303/1: 39-87.
Temporal distribution of the last hipparions (from Van der Made et al., 2022).
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
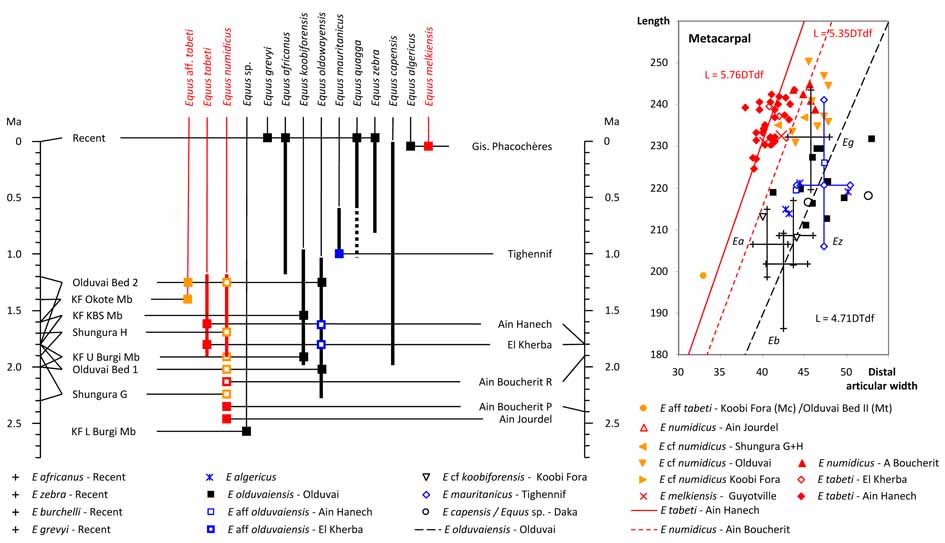
The temporal distribution of the African species of Equus (Sahnouni et al., 2018).
- Made, J. van der Made, K. Boulaghraief, R. Chelli- Cheheb, I. Cáceres, Z. Harichane & M. Sahnouni, 2022. The last North African hipparions – hipparion decline and extinction follows a common pattern. Neues Jahrbuch fürGeologie und.Paläontologie, Abhandlungen, 303/1: 39-87.
- Sahnouni, M., J.M. Parés, M. Duval, I. Cáceres, Z. Harichane, J. van der Made, A. Pérez-González, S. Abdessadok, N. Kandi, A. Derradji, M. Medig, K. Boulaghraif & S. Semaw, 2018. 1.9-million- and 2.4-million-year-old artifacts and stone tool–cutmarked bones from Ain Boucherit, Algeria. Science, 362, 1297–1301. Supplementary Materials.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
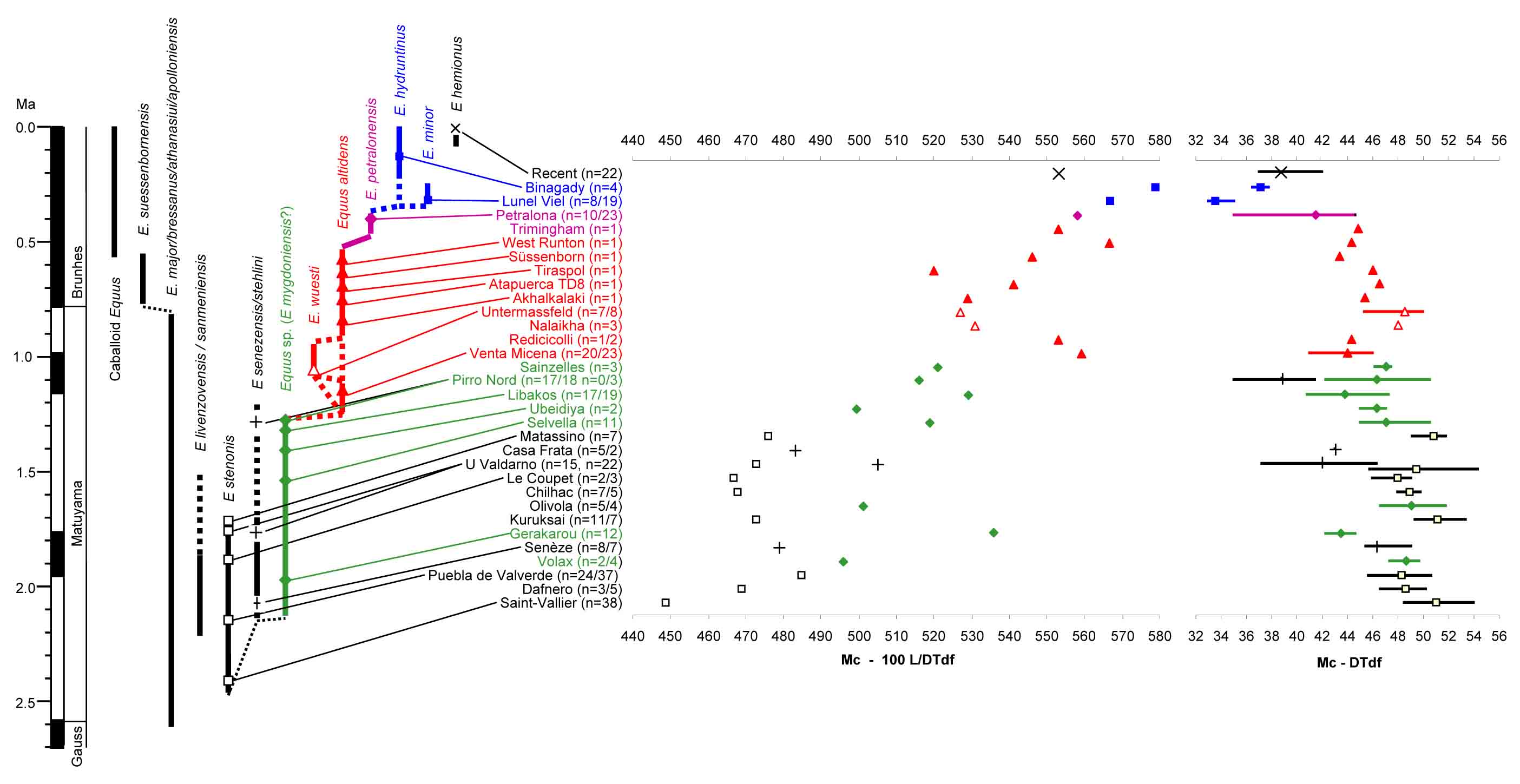
Biometrics of the metacarpal of the Equus altidens - E. petralonensis - E. hydruntinus lineage compared to other species (Van der Made et al., 2017).
- Azadbakht, S., A. Fazelioladi, N. Hashemi & J. van der Made, 2017. Gavkosh – a new locality yielding large mammal fossils in the Bakhtiari Formation in Lorestan Province, Iran. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 283/2: 151-159.
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364.
- Made, J. van der, 2013. First description of the large mammals from the locality of Penal, and updated faunal lists for the Atapuerca ungulates - Equus altidens, Bison and human dispersal into Western Europe. Quaternary International, 295: 36-47.
- Giles Pacheco, F., A. Santiago, J.M. Gutiérrez, J.M. López-García, H.-A. Blain, G. Cuenca-Bescós, J. van der Made, I. Cáceres d & N. García, 2011. The paleontological site in the Sierra del Chaparral (Villaluenga del Rosario, Cádiz-Spain). Quaternary International, 243: 92-104.
- Sánchez, A., S. Fraile, J. van der Made, J. Morales, V. Quiralte, M.J. Salesa, I.M. Sánchez, B. Sanchiz, D. Soria, J. Jiménez, L.J. Barbadillo, C. Laplana, Z. Szyndlar, 2005. Primeros datos faunísticos del Neolítico madrileño: la cueva de la Ventana (Torrelaguna, Madrid). En P.Arias Cabal, R. Ontañón Peredo, C. García-Moncó Piñeiro (eds.) III Congreso del Neolítico en la Península Ibérica. Monografias del Instituto Internacional de Investigaciones Prehistóricas de Cantabria, 1: 155-165.
- Made, J. van der, 2001. Les ongulés d'Atapuerca. Stratigraphie et biogéographie. L'Anthropologie, 105 (1): 95-113.
- Made, J. van der, 1999. Ungulates from Atapuerca-TD6. Journal of Human Evolution, 37(3-4): 389-413.
- Made, J. van der, 1998. Ungulates from Gran Dolina (Atapuerca, Burgos, Spain). Quaternaire, 9(4): 267-281.
Tapiridae
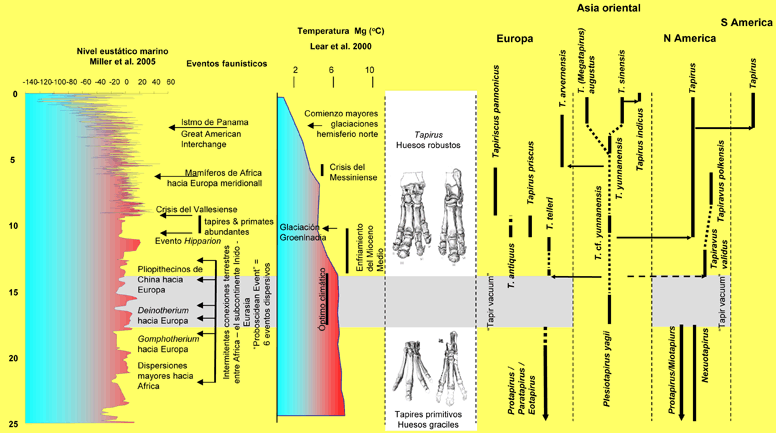
The distribution of the Neogene and Quaternary Tapiridae (from Van der Made, 2010).
- Gómez de Soler, B., G. Campeny Vall-Llosera, J. van der Made, O. Oms, J. Agustí, R. Sala, H.A. Blain, F. Burjachs, J. Claude, S. García Catalán, D. Riba1 & R. Rosillo, 2012. The Camp dels Ninots (NE Spain): a new key locality for the Middle Pliocene terrestrial faunas of Europe. Geologica Acta, 10(2): 1-17.
- Made, J. van der, 2010. Els macrovertebrats del Camp dels Ninots i el seu context: canvis ambientals, evolució i estructura social. En: G. Campeny Vall-llosera & B. Gómez de Soler (eds) El Camp dels Ninots - restres de l’evolució. Ayuntament de Caldes de Malavella, Caldes de Malavella, & Institut Català de Paleoecologia Humana i Evolució Social, Tarragona: 105-128.
- Made, J. van der & I. Stefanović, 2006. A small tapir from the Turolian of Kreka (Bosnia) and a discussion on the biogeography and stratigraphy of the Neogene tapirs. Neues Jahrbuch für Geologie und Paläontologie, Monatsheften, 240(2): 207-240.
Tubulidentata
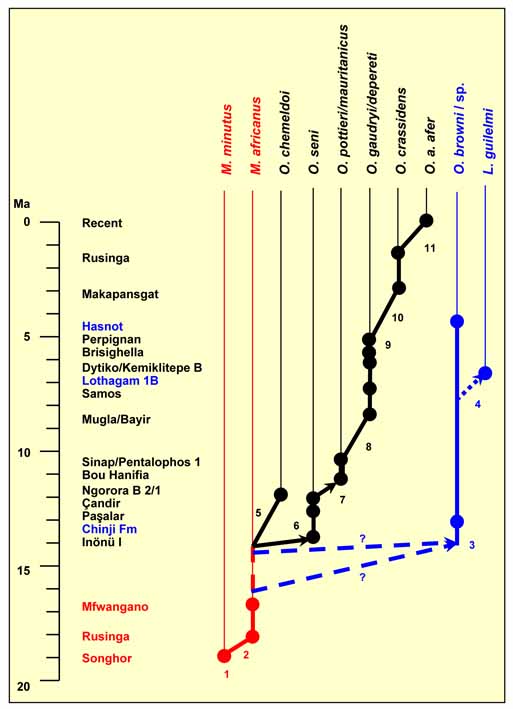
The phylogeny of the Tubulidentata (Van der Made, 2003).
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364.
- Made, J. van der, 2005. Errata and reply to Guest Editor’s notes. Courier Forschungs-Institut Senckenberg, 254: 473-477.
- Made, J. van der, 2003. The aardvark from the Miocene hominoid locality of Çandir, Turkey. Courier Forschungs-Institut Senckenberg, 240: 133-147.
Proboscidea
- Piñero, P., J. Agustí, O. Oms, I. Fierro, P. Montoya, S. Mansino, F. Ruiz-Sánchez, D.M. Alba, M.T. Alberdi, H.-A. Blain, C. Laplana, J. van der Made, A.V. Mazo, J. Morales, X. Murelaga, A. Pérez-García, F. Pérez-Valera, J.A. Pérez-Valera, P. Sevilla, J.M. Soria, G. Romero, 2017. Early Pliocene continental vertebrate fauna at Puerto de la Cadena (SE Spain) and its relevelance to the marine-continental correlation of the Late Neogene of Eastern Betics. Palaeogeography, Palaeoclimatology, Palaeoecology, 479: 102–114.
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364.
- Made, J. van der, 2014. La evolución de los macromamíferos africanos del Plio-Pleistoceno. / The Plio-Pleistocene large mammals of Africa - Why they evolved like they did. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 137-177, English 308-323, references 360-362.

The transition of Gomphoterium to Tetralophodon in the Iberian Peninsula (Mazo & Van der Made, 2012).
- Mazo, A.V. & J. van der Made, 2012. Iberian mastodonts: geographic and stratigraphic distribution. Quaternary International, 255: 239-256.
- Made, J. van der, 2010. The evolution of the elephants and their relatives in the context of changing climate and geography. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 340-360.
- Mazo, A.V. & J. van der Made, 2010. Iberian mastodonts: geographic and stratigraphic distribution. Quaternaire, Horse série 3: 170.
- Stefanovic, I., A.V. Mazo & J. van der Made, 2010. Geographic and stratigraphic distribution of the Serbian mastodonts and deinotheres. Quaternaire, Horse série 3: 172-173.
- Made, J. van der & A.V. Mazo, 2003. Proboscidean dispersals from Africa towards western Europe. Deinsea, 9: 437-452.
- Made, J. van der & A.V. Mazo, 2001. Spanish Pleistocene proboscidean diversity as a function of climate. En: (G. Cavarretta, P. Gioia, M. Mussi, M.R. Palombo, eds.) The world of elephants. Proceedings of the 1st international congress. Consiglio Nazionale delle Ricerche, Roma: 214-218.
Primates
- Ramírez-Pedraza, I., L.M. Martínez, H. Aouraghe, F. Rivals, C. Tornero, H. Haddoumi, F. Estebaranz-Sánchez, A. Rodríguez-Hidalgo, J. van der Made, A. Oujaa, J.J. Ibáñez, H. Mhamdi, M. Souhir, A. Al Mahdi, M.G. Chacón & R. Sala-Ramos, 2023. Multiproxy approach to reconstruct fossil primate feeding behavior: Case study for macaque from the Plio-Pleistocene site Guefaït-4.2 (eastern Morocco). Frontiers in Ecology and Evolution, 11:1011208.
- Alba, D.M., A. Rodríguez-Hidalgo, H. Aouraghe, J. van der Made, A. Oujaa, H. Haddoumi, P. Saladié, A. Al Mahdi, J. Marín, M. Farkouch, C. Lorenzo, S. Bengamra, E. Delson, M.G. Chacón & R. Sala-Ramos, 2021. New macaque fossil remains from Morocco. Journal of Human Evolution, 153: 102951.
- Made, J. van der, 2014. Late Pleistocene European and Late Miocene African acceleration of faunal change in relation to climate and as a background to human evolution. Quaternary International, 326-327: 431-447.
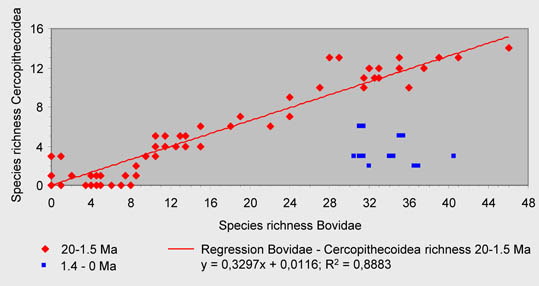
Species richness of Bovidae and Cercopithecoidea in Africa. From Van der Made, 2014.
- Harrison, T, J. van der Made & F. Ribot, 2002. A pliopithecin from Sant Quirze (Middle Miocene, Spain) and remarks on pliopithecid biochronology and zoogeography. Journal of Human Evolution, 42(4): 371-377.
Right M2 from Hostalets (A) and San Quirze (B), the first assigned to Dryopithecus and the second to Pliopithecus (Van der Made & Ribot, 1999; Harrison et al., 2002). These specimens were collected in the 1920-ties by the aficionado M. Guérin, later they passed on to the Museu i Laboratori de Geologia del Seminari in Barcelona. When studying the fossil pigs from these localities in 1991, I recognized that these specimens are no Suidae, but primates. These are the first Miocene primate fossils found in Spain after a Dryopithecus specimen Seu d'Urgell, described y Vidal in 1913, and are the first from the Vallès-Penedès Basin. At present, this basin is where most Miocene primates from Europe have been found.
- Made, J. van der & F.Ribot, 1999. Additional hominoid material from the Miocene of Spain and remarks on hominoid dispersals into Europe. Contributions to Tertiary and Quaternary Geology, 36(1-4): 25-39.
Carnivora
- Walker, M.J., M. Haber Uriarte, A. López Jiménez, M. López Martínez, J. van der Made, M. Duval, R. Grün & I. Martín Lerma, 2020. Cueva Negra del Estrecho del Río Quípar: A dated late Early Pleistocene Palaeolithic site in southeastern Spain. Journal of Paleolithic Archaeology, 3: 816–855.
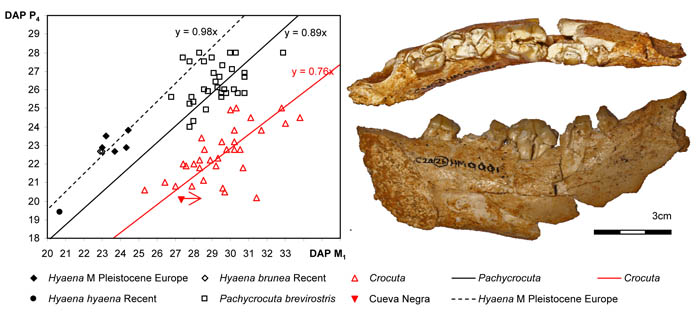
Crocuta from Cueva Negra, one of the oldest occurrences in Europe (Walker et al., 2020).
- Vaquero, M., J. van der Made, H.A. Blain, N. Ibáñez, J.M. López-García, F. Rivals, S. Alonso, A. Ameijenda, M. Bennàsar, M. Fernandez-García, A. de Lombera & I. Valverde, 2018. Fauna, environment and human presence during MIS5 in the North of Spain: The new site of Valdavara 3. Comptes Rendus Palevol, 17/8: 557-593. Appendix A. Supplementary data.
- Made, J. van der, T. Torres, J.E. Ortiz, L. Moreno-Pérez & Y. Fernández Jalvo, 2016. The new material of large mammals from Azokh and comments on the older collections. In: Fernández-Jalvo, Y., T. King, P. Andrews & L. Yepiskoposyan (eds). Azokh caves and the transcaucasian corridor. Springer - Dordrecht: 117-162.
- Made, J. van der & A.V. Mazo, 2014. Los grandes mamíferos del yacimiento de Preresa. En: I. Baquedano Beltrán & C. Laplana (eds) Haploidoceros mediterraneus una nueva especie de ciervo en el Pleistoceno Ibérico. Museo Arqueológico Regional - Alcalá de Henares: 39-53.
- Made, J. van der, K. Stefaniak & A. Marciszak, 2014. The Polish fossil record of the wolf Canis and the deer Alces, Capreolus, Megaloceros, Dama and Cervus in an evolutionary perspective. Quaternary International, 326-327: 406-430.
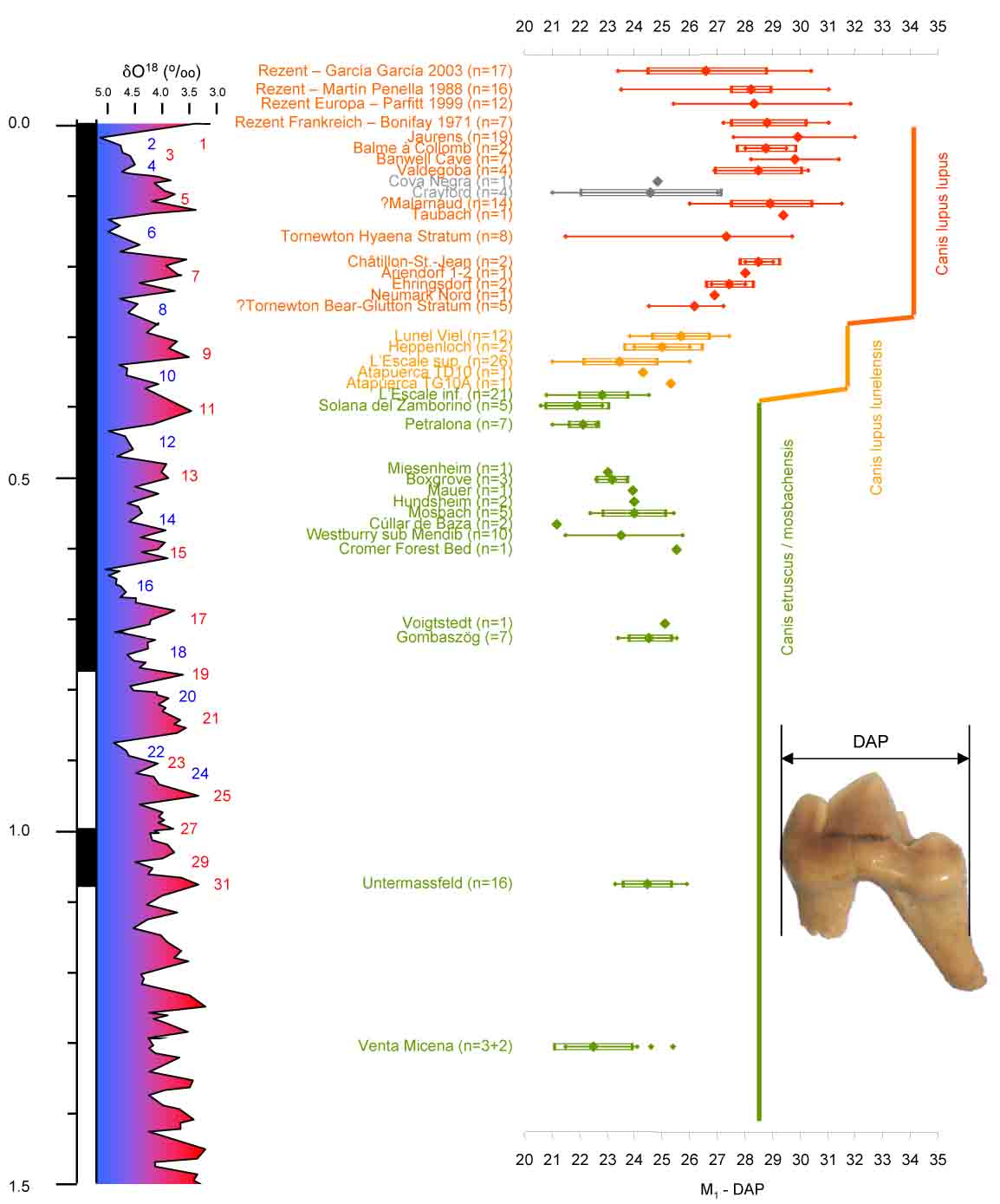
The size increase in the Wolf lineage as seen in the increase in length of the lower carnassial (Van der Made, 2010).
- Made, J. van der, 2010. Biostratigraphy - "Large Mammals". In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 82-92.
- Varela, S., J.M. Lobo, J. Rodríguez & J. van der Made, 2010. ¿Las extinciones de herbívoros a finales del Pleistoceno están relacionadas con la desaparición de las hienas manchadas? Zona Arqueológica, 13: 76- 89.
- Canals, A., J. van der Made, I. Sauceda & E. Carbonell, 2003. El conjunto paleontológico de la Cueva de Maltravieso (Cáceres, Spain): un nuevo yacimiento del Pleistoceno. XI Reunión Nacional de Cuaternario. Oviedo, 2-4/7/2003: 313-320.
- Made, J. van der, 1999. Biogeography and stratigraphy of the Mio-Pleistocene mammals of Sardinia and the description of some fossils. Deinsea, 7: 337-360.
- Mazo, A.V., J. van der Made, J.F. Jordá, E. Herraez, & I. Armenteros, 1998. Fauna y bioestratrigrafía del yacimiento Aragoniense de Montejo de la Vega de la Serrezuela (Segovia). Estudios Geológicos, 54(5-6): 231-248.
3) Biogeography, biostratigraphy, "the history of nature", extinction
- Made, J. van der Made, K. Boulaghraief, R. Chelli- Cheheb, I. Cáceres, Z. Harichane & M. Sahnouni, 2022. The last North African hipparions – hipparion decline and extinction follows a common pattern. Neues Jahrbuch fürGeologie und.Paläontologie, Abhandlungen, 303/1: 39-87.
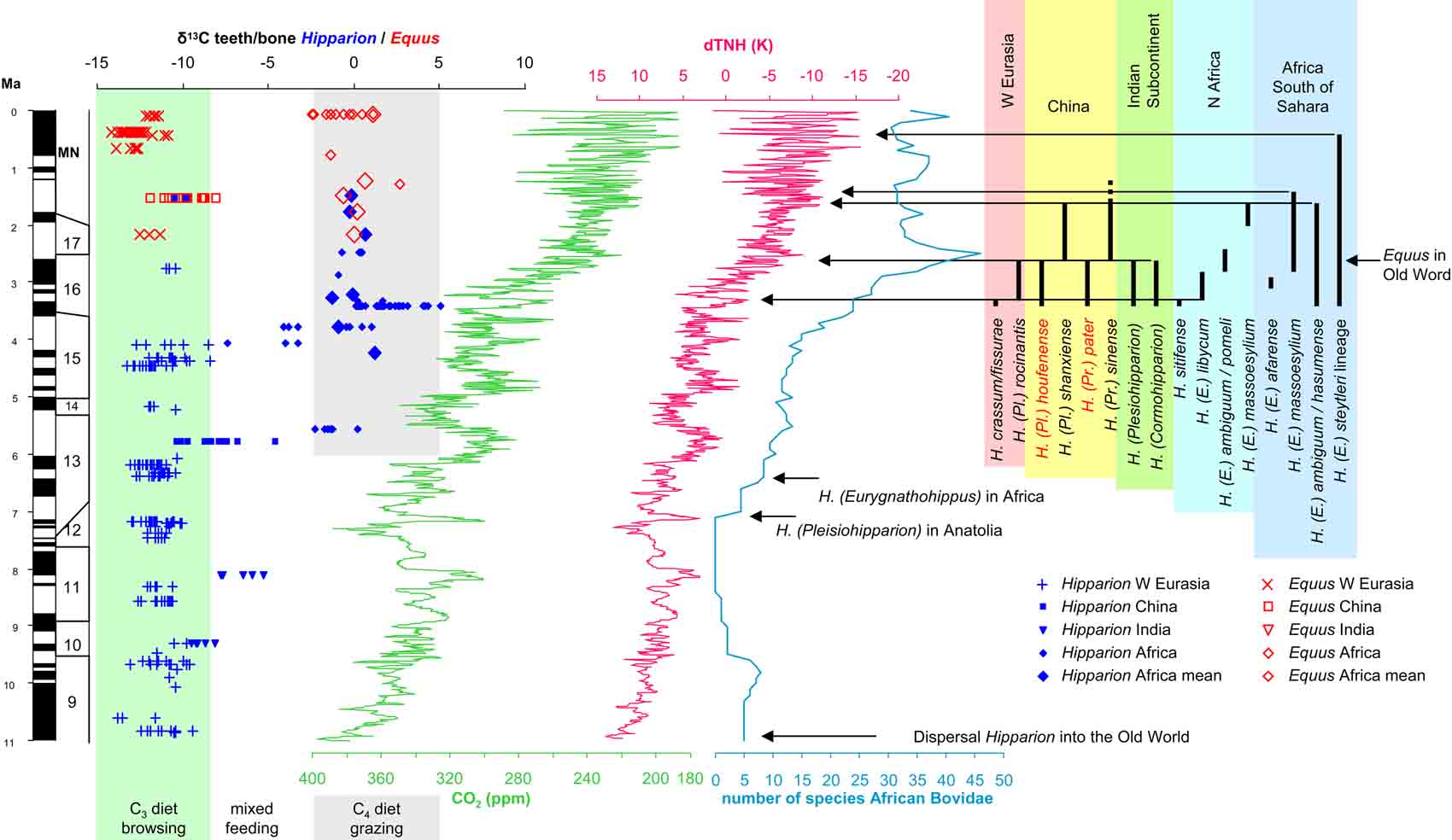
The last hipparions started to go extinct after about 2.5 Ma. This happened first in the North, while near to the equator they remained diverse and went extinct much later. A similar pattern of extinction is observed in many other groups. The radiation of the Bovidae in Africa does not seem to have been at the cost of the hipparions. The decline and extction of the hipparions happened against a backfround of decreasing atmospheric CO2 and global temperature. There is no evidence that the spread of C4 grasses (which is related to decreasing CO2) affected the hipparions. because, like Equus, they fed on the available vegetation: C4 grasses near the equator and C3 plants at higher latitudes. (From Van der Made et al., 2022.)

Chronostratigraphic chart of the Quaternary (Silva et al., 2021).
- Silva, P.G., T. Bardají, J. Baena-Preysler, J.L. Giner-Robles, J. van der Made, C. Zazo, A. Rosas & J. Lario, 2021. Tabla cronoestratigráfica del Cuaternario de la península ibérica (v 3.0): Nuevos datos estratigráficos, paleontológicos y arqueológicos. Cuaternario y Geomorfología, 35(3-4): 121-146.
- Silva, P.G., T. Bardají, J. Baena, J.L. Giner-Robles, J. van der Made, C. Zazo, A. Rosas & J. Lario, 2021. Tabla Cronoestratigráfica del Cuaternario de la Península Ibérica v 3.0 Chronostratigraphic Chart for the Quatemary of the Iberian Peninsula v 3.0. Geo-Temas, 18; 1035, la tabla en la contraportada.
- Made, J. van der, 2021. Las extinciones en el pasado, previsión para el futuro. In: C. Cánovas Fernández (ed.). Museo Nacional de Ciencias Naturales. Nuestra investigación al alcance de tu mano. Consejo Superior de Investigaciones Científicas - Madrid: 378-383.
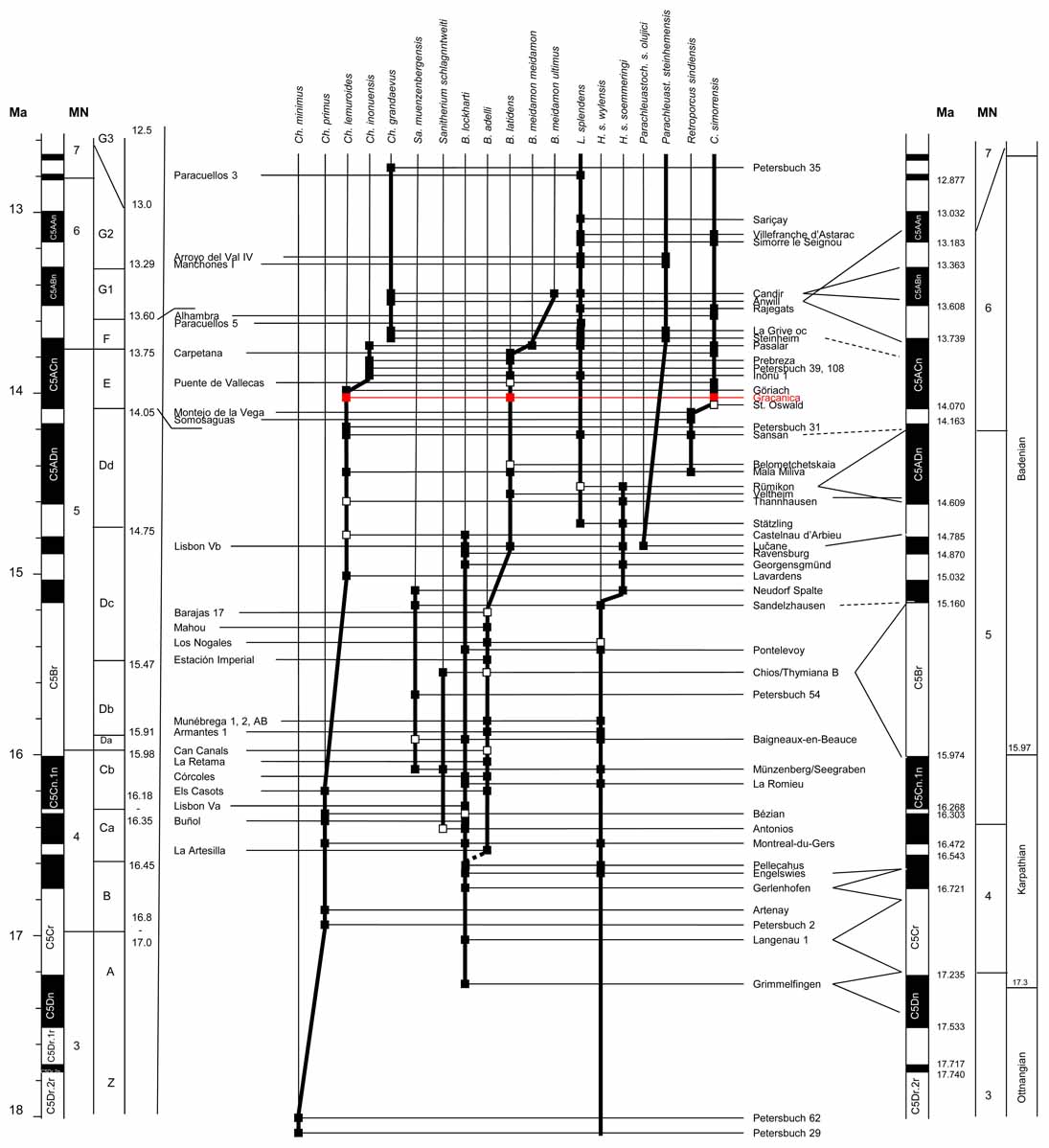
Suoidea and correlation (Van der Made, 2020). The genus Choeromorus is indicated on the left and the grades of evolution are well seen in the length of the M3 (see below). There are similar changes in other lineages (see also Bunolistriodon in the Suoidea section). Using the evolution of different lineages, the localities can be ordered in an age sequence. MN units, as they are currently applied, appear to be diachronic: compare colums on the left (Spain) and right (Germany and Austria). The grades of evolution allow a correlation from Spain across Europe to Anatolia and, unlike the MN units, are not in contradiction with other means to estimate the ages of the localities.
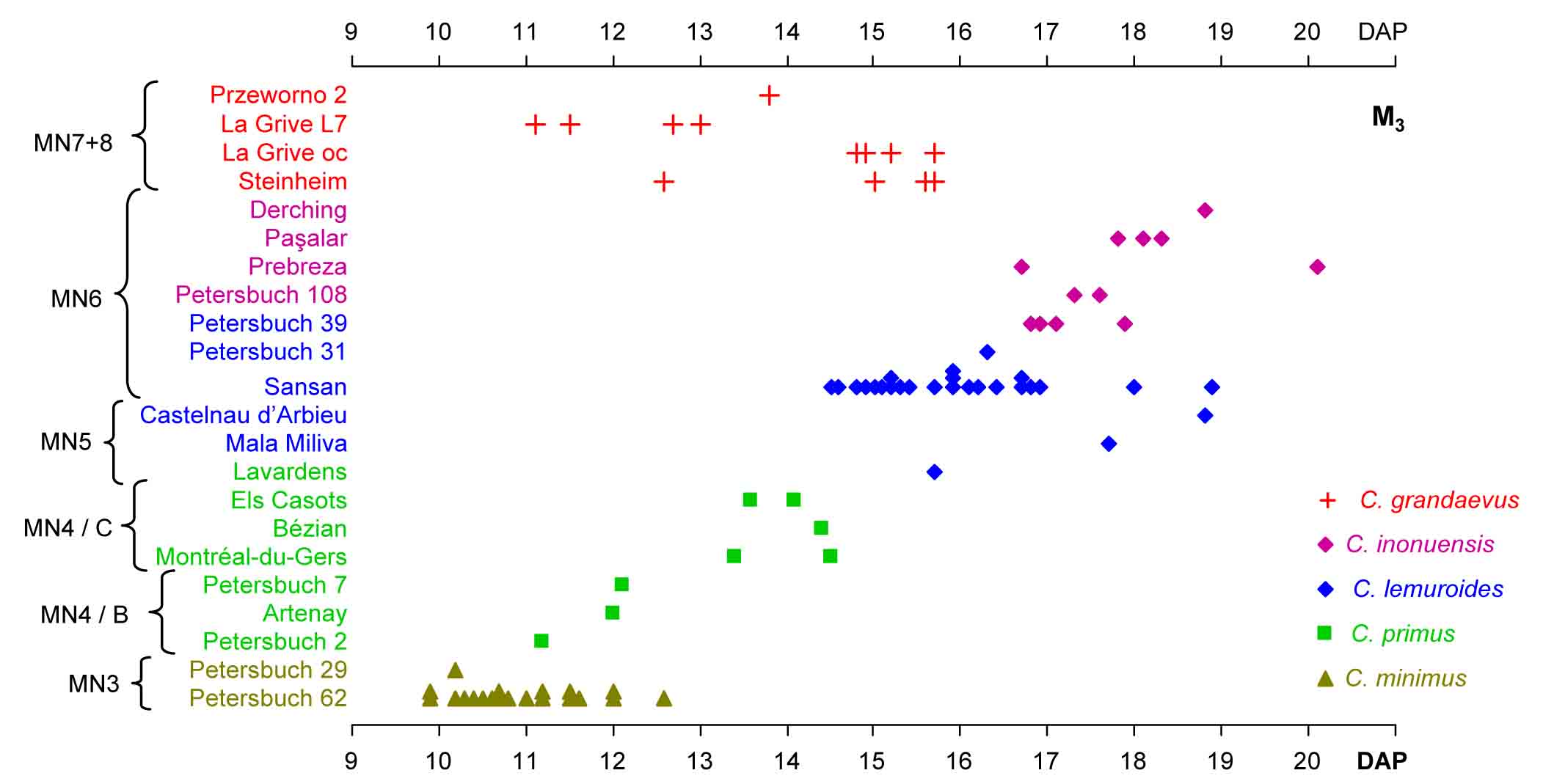
The increase in length (DAP) in the M3 in Choeromorus (= Taucanamo) (Van der Made, 2020). The first four species form an anagenetic lineage, the fifth species is an offshoot of this lineage. There are also morphological changes, but the increase in length is a very clear trend that can described by measurements.
- Made, J. van der, 2020. The Suoidea from the Middle Miocene of Graćanica (Bugojno Basin, Bosnia, and Herzegovina) — evolution, taxonomy, and biostratigraphy. Palaeobiodiversity and Palaeoenvironments, 100: 321–349. Electronic supplementary material.
- Made, J. van der, 2019. The dwarfed "giant deer" Megaloceros matritensis n.sp. from the Middle Pleistocene of Madrid - a descendant of M. savini and contemporary to M. giganteus. Quaternary International, 520: 110-139.
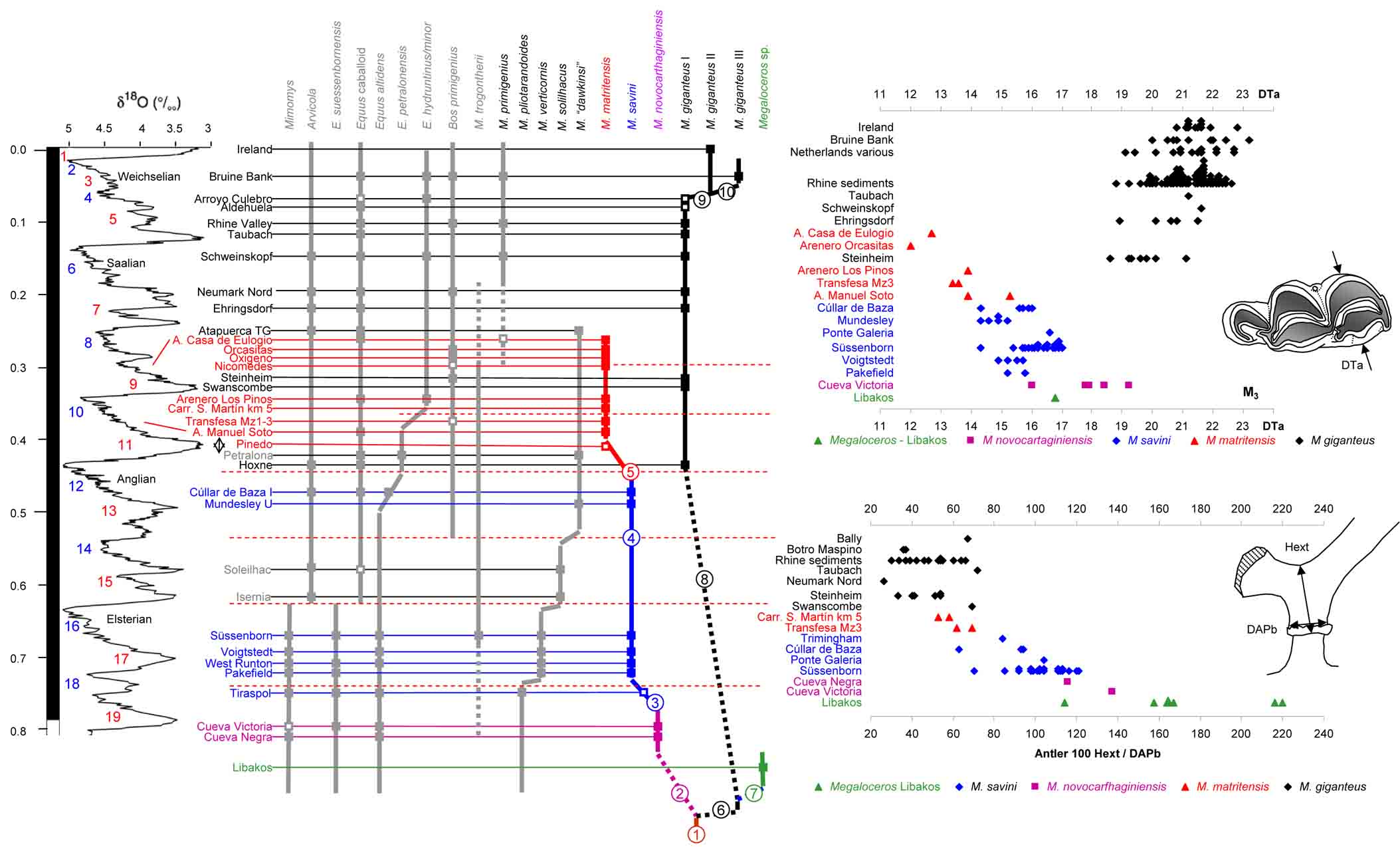
Material previously assigned to Megaloceros savini led to an overestimation of the ages of the terraces of the Mananares river, which became apparent when ESR dates for these terraces became available. The recognition of these fossils as a distinct species, Megaloceros matritensis, solved the problem (Van der Made, 2019). The species is a descendant of M. savini and differs in many characters, including smaller size (as indicated by the width of the first lobe of the M3) and a lower bifurcation between the brow tine and main beam (as indicated by the index 100Hext/DAPb).
- Made, J. van der, 2018. Quaternary large mammalian zones. En: S.L. López Varela (ed.) The Encyclopedia of Archaeological Sciences. JohnWiley & Sons, Inc.: 1428-1431.
- Made, J. van der, J. Rosell & R. Blasco, 2017. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Quaternary International, 433: 296-346.
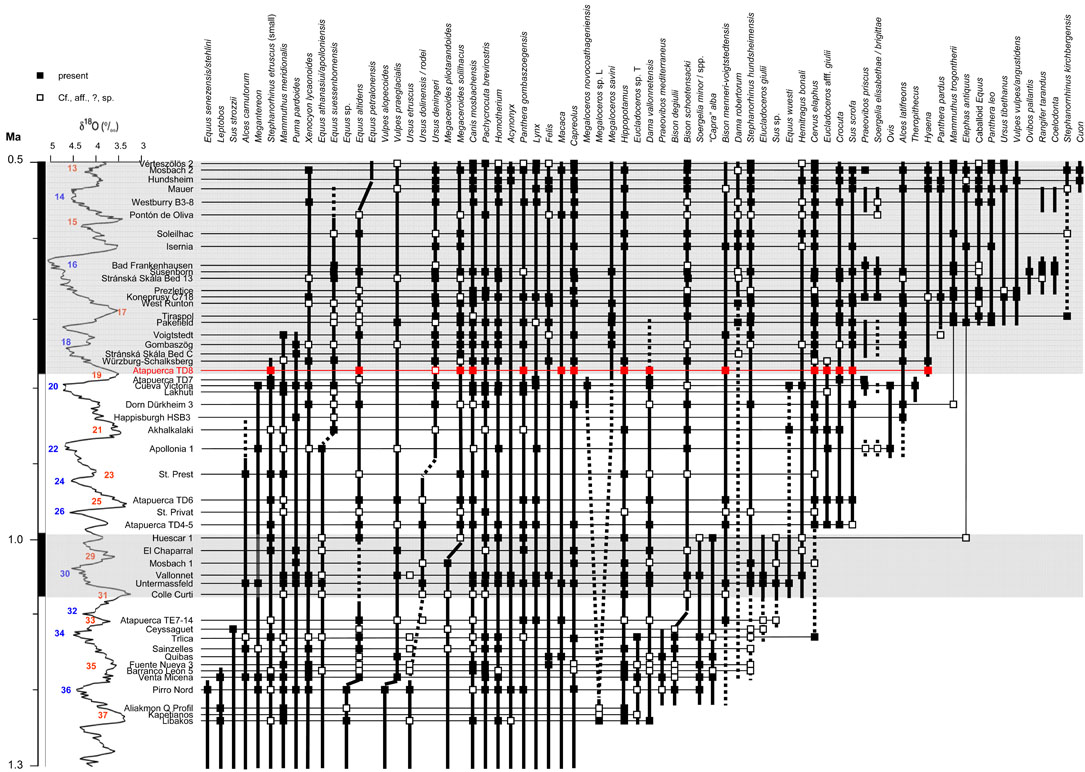
The stratigraphic distirbution of the large mammals around the Early-Middle Pleistocene boundary (Van der Made et al., 2017).
- Made, J. van der, M. Sahnouni & Kamel Boulaghraief, 2017. Hippopotamus gorgops from El Kherba (Algeria) and the context of its biogeography. In: M. Sahnouni, S. Semaw & J. Rios Garaizar (eds.) Proceedings of the II Meeting of African Prehistory, Burgos 15-16 April, 2015. Centro Nacinonal de Investigación sobre la Evolución Humana - Burgos: 135-169.
- Kirilovaa, I.V., O.F. Chernova, J. van der Made, V.V. Kukarskih, B. Shapiro, J. van der Plicht, F.K. Shidlovskiya, P.D. Heintzman, T. van Kolfschoten, O.G. Zanina, 2017. On the discovery of the skull of Stephanorhinus kirchbergensis (Jäger, 1839) above the Arctic Circle. Quaternary Research, 88/3: 537-550.
- Kirilovaa, I.V., O.F. Chernova, J. van der Made, V.V. Kukarskih, B. Shapiro, J. van der Plicht, F.K. Shidlovskiya, P.D. Heintzman, T. van Kolfschoten, O.G. Zanina, 2017. Discovery of the skull of Stephanorhinus kirchbergensis (Jäger, 1839) above the Arctic Circle – CORRIGENDUM. Quaternary Research, 88: 570.
- Made, J. van der, 2014. Late Pleistocene European and Late Miocene African acceleration of faunal change in relation to climate and as a background to human evolution. Quaternary International, 326-327: 431-447.
- Made, J. van der, 2014. Los grandes mamíferos del Plio-Pleistoceno africano: Afrotheria, Perissodactyla y Artiodactyla I. / The large mammals of the Plio-Pleistocene of Africa: Afrotheria, Perissodactyla and Artiodactyla I. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 179-215, English 324-336, references 362-364.
- Made, J. van der, 2014. La evolución de los macromamíferos africanos del Plio-Pleistoceno. / The Plio-Pleistocene large mammals of Africa - Why they evolved like they did. In: M. Dominguez Rodrígo & E. Baquedano (eds.) La Cuna de la Humanidad. Museo Arquelógico Regional, Alcalá de Henares & Museo de la Evolución Humana, Burgos: 137-177, English 308-323, references 360-362.
- Made, J. van der, 2013. First description of the large mammals from the locality of Penal, and updated faunal lists for the Atapuerca ungulates - Equus altidens, Bison and human dispersal into Western Europe. Quaternary International, 295: 36-47.
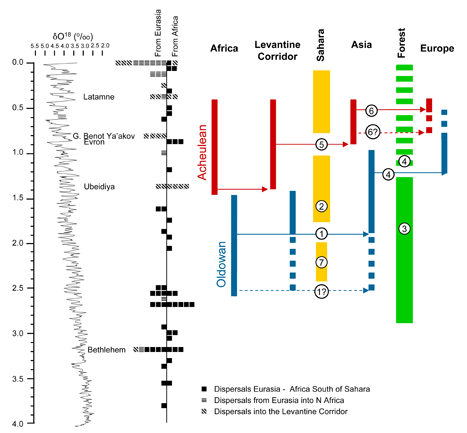
Human dispersal from Africa into Eurasia. Adapted from Van der Made (2013). The arid belt that runs through the Sahara and Middle East and the forested area of central Europe have been barriers for many species, including humans, during much of the time, but could be crossed at particular times. The dispesals of large mammals have been used to determine the moments when these barriers could be crossed. This model provides an explanation for the paradox that the humans with primitive Oldowan technology dispersed first in Europe and not those using the Achelean, which would be expected to be better colonizers as they had a more advanced technology.
- Made, J. van der, 2013. Faunal exchanges through the Levantine Corridor and human dispersal: The paradox of the latedispersal of the Acheulean industry. In: M. Sahnouni (ed.) Proceedings of the International Symposium Africa, Cradle Of Humanity: Recent Discoveries. Travaux du Centre National de Recherches Préhistoriques, Anthropologiques et Historiques, Nouvelle série 18: 255-296.
- Made, J. van der, 2013. Clima, fauna y dispersión humana desde África a Eurasia. Periodico de Atapuerca, 25: 2.
- Made, J. van der, 2011. Biogeography and climatic change as a context to human dispersal out of Africa and within Eurasia. Quaternary Science Reviews, 30: 1353-1367.
- Made, J. van der & J. Morales, 2011. Mitilanotherium inexpectatum (Giraffidae, Mammalia) from Huélago (Lower Pleistocene; Guadix-Baza basin, Granada, Spain) - observations on a peculiar biogegraphic pattern. Estudios geológicos, 67(2): 613-627.
- Made, J. van der, 2010. The evolution of the elephants and their relatives in the context of changing climate and geography. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 340-360.
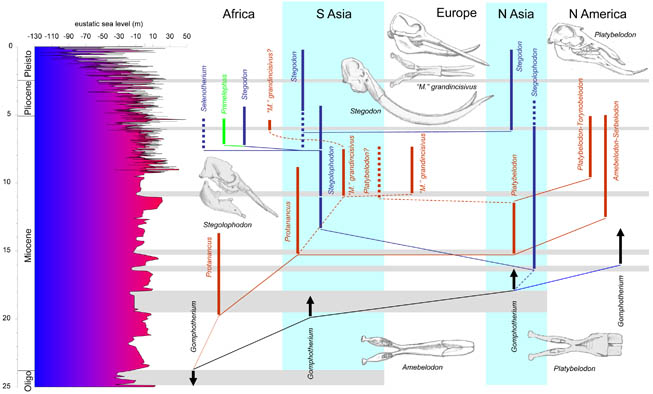
The evolution and biogeography of the gomphotheres and allies (Van der Made, 2010.)
- Made, J. van der, 2010. Biostratigraphy - "Large Mammals". In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 82-92.
- Made, J. van der, 2010. Megafauna extinctions in the wake of human dispersal. In: D. Höhne & W. Schwarz (eds) "Elefantentreich - Eine Fossilwelt in Europa". Landesamt für Denkmalpflege und Archälogie Sachsen-Anhalt & Landesmuseum für Vorgeschichte, Halle: 592-606.
- Made, J. van der & A. Mateos, 2010. Longstanding biogeographic patterns and the dispersal of early Homo out of Africa and into Europe. Quaternary International, 223-224: 195-200.
- Made, J. van der, J. Morales & P. Montoya, 2006. Late Miocene turnover in the Spanish mammal record in a wider context. Palaeogeography, Palaeoclimatology, Palaeoecology, 238: 228-246.
- Made, J. van der, 2005. Considerations on dispersals between Africa and Europe across the Strait of Gibraltar. In: J. Rodríguez Vidal,c. Finlayson & F. Giles Pacheco (eds.). Cuaternario Mediterraneo y poblamiento de hominidos. AEQUA & Gibraltar Museum, Gibraltar: 91-92.
- Made, J. van der, 2005. El Clima y su funcionamiento. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 1 Introducción, Sección 1.3 El entorno natural. Ariel - Barcelona: 37-44.
- Made, J. van der, 2005. La fauna del Plio-Pleistoceno. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Cápitulo 2 - África, Sección 2.3. Ariel, Barcelona: 71-102.
- Made, J. van der, 2005. La fauna. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 3 - Asia; Sección 3.4. Ariel, Barcelona: 270-306.
- Made, J. van der, 2005. La fauna del Pleistoceno europeo. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 4 - Europa; Sección 4.4. Ariel: 394-432.
- Made, J. van der, 2005. La fauna de Australia y Nueva Guinea. La extinción de la megafauna. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 5 - Oceanía, Sección 5.5. Ariel, Barcelona: 578-582.
- Made, J. van der, 2005. La fauna americana. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 6 - América; Sección 6.2. Ariel: 611-625.
- Made, J. van der, 2005. Las extinciones del Pleistoceno-Holoceno. Cambio climátio o acción antrópica. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 4 - Europa; Sección 4.24. Ariel, Barcelona: 554-555.
- Made, J. van der, 2001. Biogeografía y colonización de Europa. La Vanguardia, lunes, 5 de noviembre, 2001: p. 29.
- Made, J. van der, 1999. Intercontinental relationship Europe-Africa and the Indian Subcontintent. In (G. Rössner & K. Heissig, eds.) The Miocene land mammals of Europe. Verlag Dr. Friedrich Pfeil, München: 457-472.
- BiochroM'97 1997. Synthèses et tableaux de corrélations. Mémoires et Travaux EPHE, Montpellier, 21: 769-805. (Biochrom'97 incluye a muchos autores, entre ellos J. van der Made.)
- Made, J. van der 1997. Intercontinental dispersal events, eustatic sea level and Early and Middle Miocene Stratigraphy. Mémoires et Travaux EPHE, Montpellier, 21: 75-81.
- Made, J. van der, 1996. Listriodontinae (Suidae, Mammalia), their evolution, systematics and distribution in time and space. Contributions to Tertiary and Quaternary Geology, 33(1-4): 3-254, 54 pp. 19 tables on a separate microfiche.
- Made, J. van der, 1996. Pre-Pleistocene land mammals from Crete. D. S. Reese (ed.): The Pleistocene and Holocene Fauna of Crete and its First Settlers. Monographs in World Archaeology, 28: 69-79.
- Made, J. van der, 1992. Migrations and climate. Courier Forschungsinstitut Senckenberg, 153, 27-39.
4) Insular ecology and evolution
As I was a student of Paul Sondaar, it is only natural that I am intersted in the evolution of insular mammals. His model is published as a book chapter and as a paper in the journal Natural History and is illustrated by the three cartoons below. In fact it is only one of the ways endemic insular faunas may originate.

Insular faunas may originate during periods of low sea level, when the distance from the main land to the island is less (from Sondaar, 1977; Sondaar et al., 1978). Only the animals which are good swimmers, like hippos, deer and elephants, arive to the islands, while worse swimmers, like horses and carnivores do not reach the islands.

Insular faunas become isolated and endemic when sea level rises and the distance from the main land to the island increases (from Sondaar, 1977; Sondaar et al., 1978). In the absence of carnivores, large mammals dwarf and small mammals become giants. Selection pressure is different and a number of adaptations evolve: low gear locomotion, seen in different proportions of the limb bones, high crowned teeth, stereo vision (eyes rotate forewards, the fields of view overlap and the animal can see depth), etc.

During the Pleistocene, ice ages became more severe and sea levels dropped deeper and deeper. When islands became connected to the main land, their endimic faunas went extinct (from Sondaar, 1977; Sondaar et al., 1978). The person to the left is Paul Sondaar.
Sondaar's work on insular faunas and ecology was original in that it goes beyond calculations of species numbers and body size changes, and shows a relationship between insular ecology and morphological adaptations. This relationship caused these adaptations to evolve convergently in islands in different parts of the world. For instance the shortened metapodials, which evolved in Mediterranean islads and in the Ryukyu Islands (see graphs below). The degree to which metapodials became shortened is related to the presence or absence of carnivores. In addition the progressively more severe glaciations led to islands becoming less isolated and their successive faunas beoming less endemic as can be seen in the range charts for the different islands below.
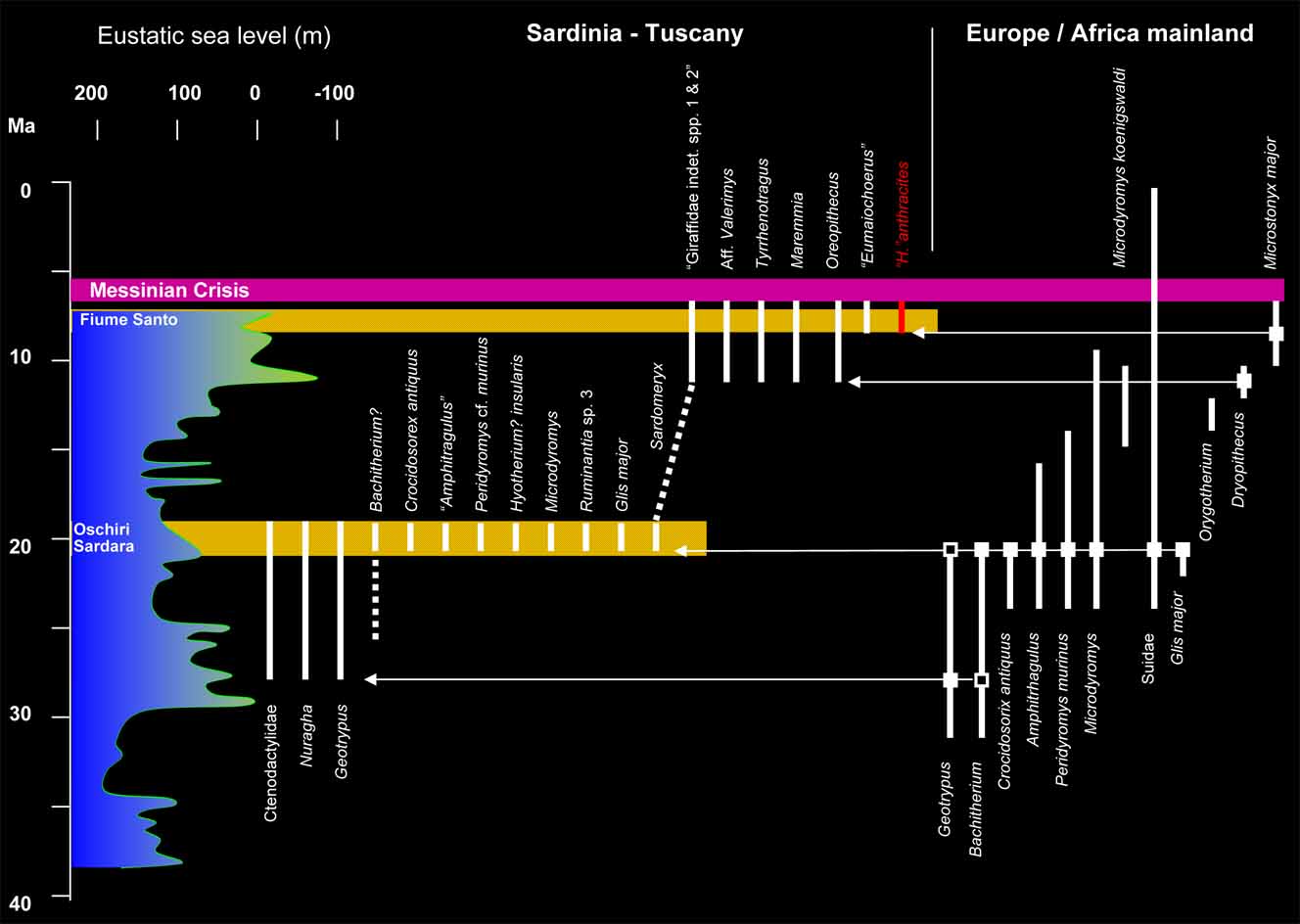
Faunal evolution of Miocene island/achipelago of Tuscany, Corsica andSardinia (Van der Made, 2008).
- Made, J. van der, 2008. New endemic large mammals from the Lower Miocene of Oschiri (Sardinia), observations on evolution in insular environment. Quaternary International, 182: 116-134.
- Made, J. van der & M.R. Palombo, 2006. Megaloceros sardus n.sp., a large deer from the Pleistocene of Sardinia. Annales Géologiques des Pays Helléniques, 41: 163-176.
- Made, J. van der, 2005. The fossil endemic goat Nesogoral cenisae n.sp. from Campidano Sardinia - cursorial adaptations in insular environment. Monografías Societat d’Història Natural de les Balears, 12: 347-368.
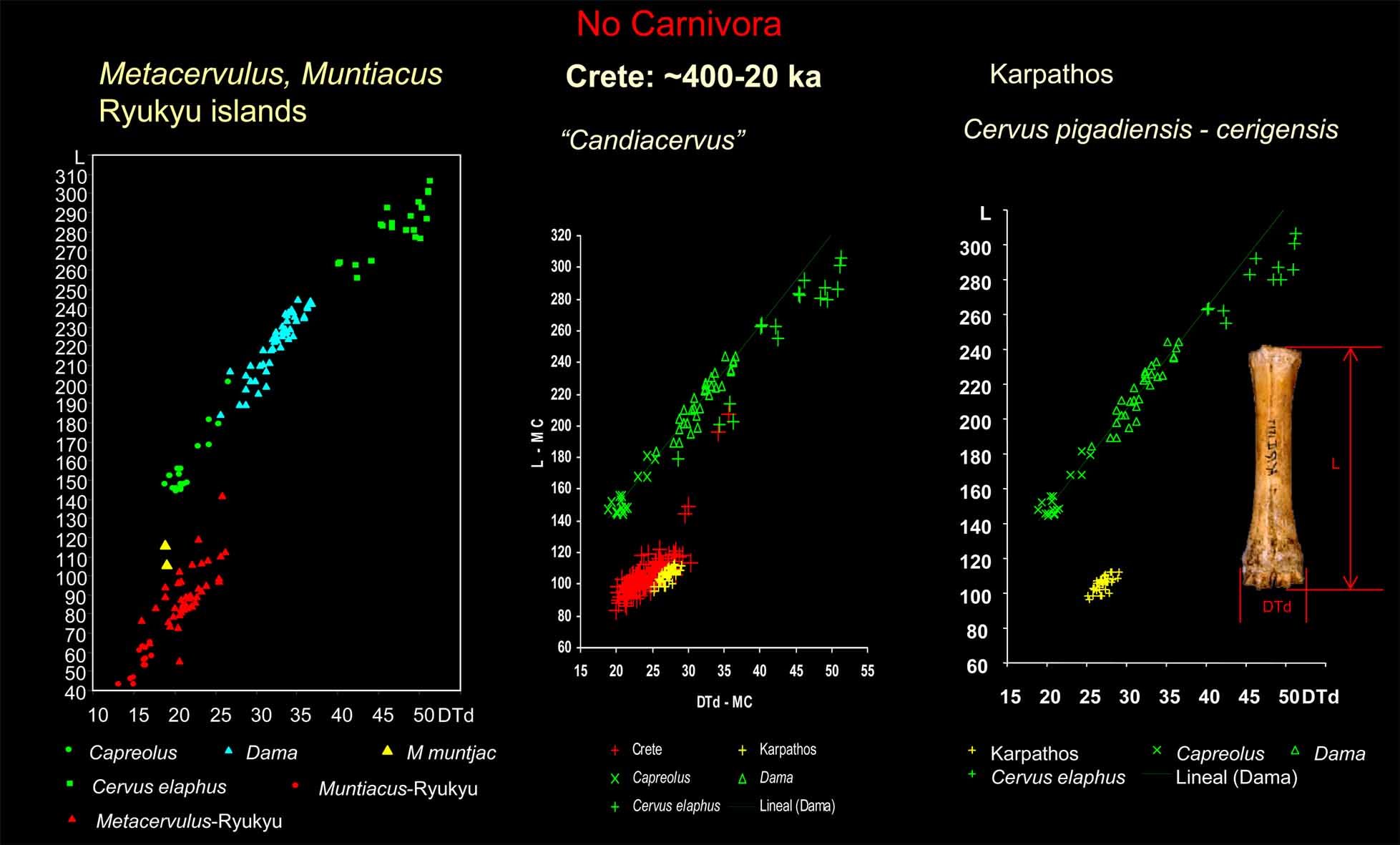
Endemic deer species from islands without carnivores, such as the Pleistocene Ryukyu Islands, Crete and Carpathos, have shortened metapodials compared to the main land species (Capreolus spp., Dama spp. Cervus elaphus) (Van der Made, 2005).
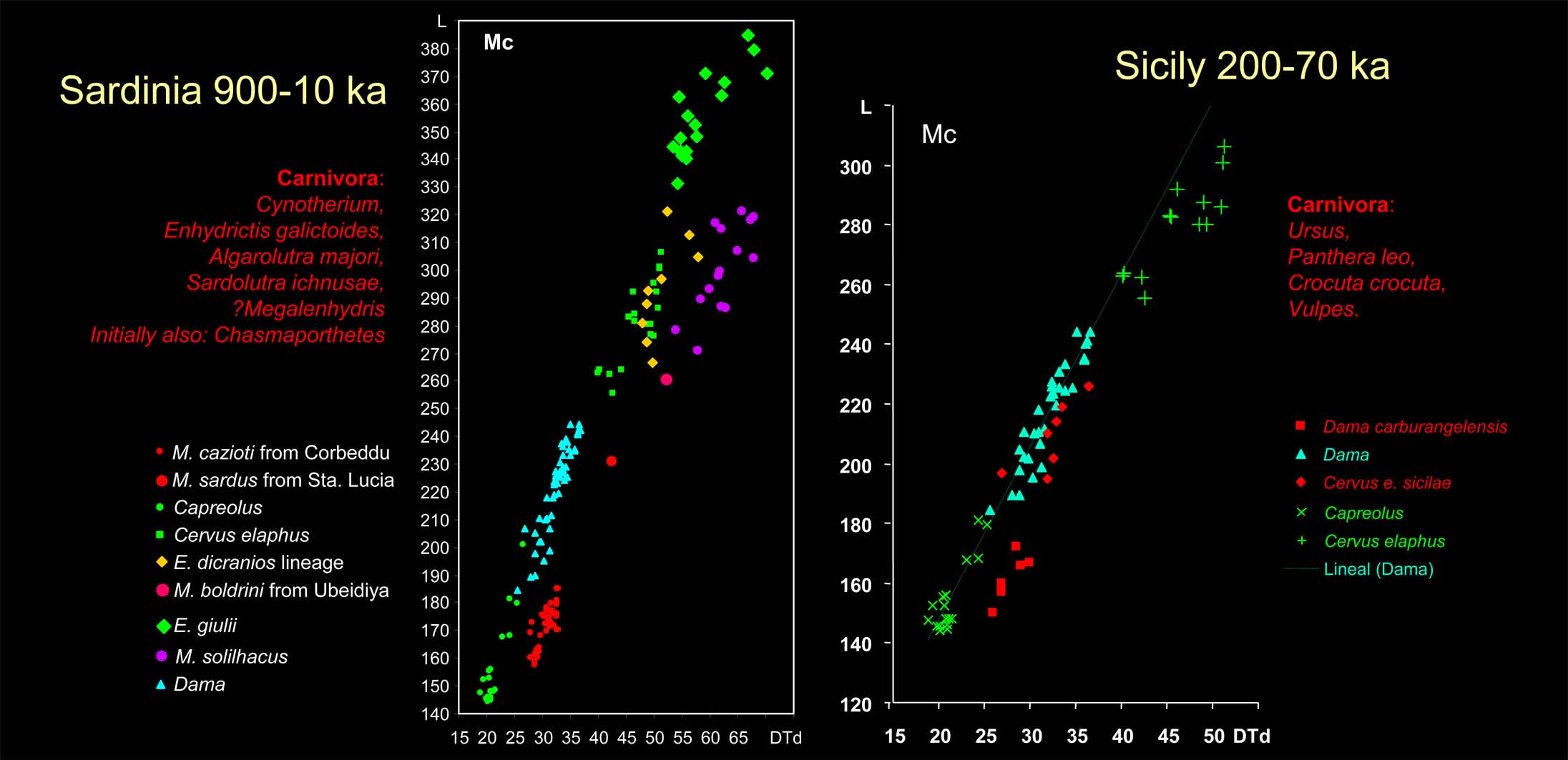
Endemic deer species from islands with carnivores, such M. cazioti, M. sardus, Dama carburangelensis and Cervus elaphus siciliae, have metapodials that are little or not shortened compared to the main land species (Capreolus spp., Dama spp., Cervus elaphus, Eucladoceros and Megaceroides) (Van der Made, 2005).
- Made, J. van der, 2005. La fauna del Plio-Pleistoceno. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Cápitulo 2 - África, Sección 2.3. Ariel, Barcelona: 71-102.
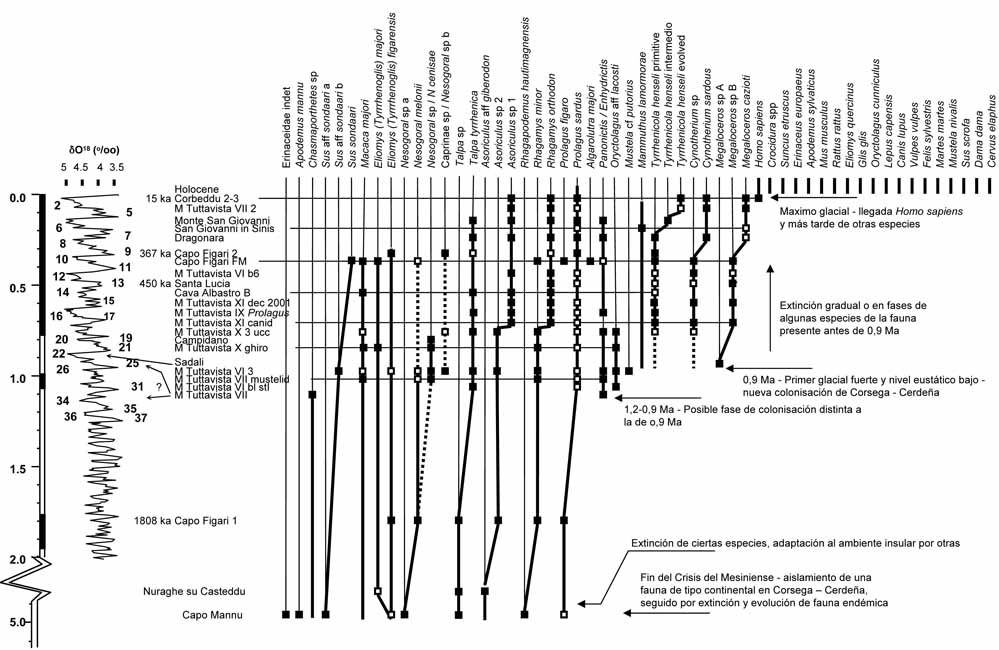
Range chart of the Pliocene and Quaternary mammals from Sardinia (Van der Made, 2005).
Range chart of the Quaternary mammals from Sicily (Van der Made, 2005).
Range chart of the Quaternary mammals from Crete (Van der Made, 2005).
- Made, J. van der, 2005. La fauna. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 3 - Asia; Sección 3.4. Ariel, Barcelona: 270-306.
Range chart of the Quaternary mammals from Java (Van der Made, 2005).
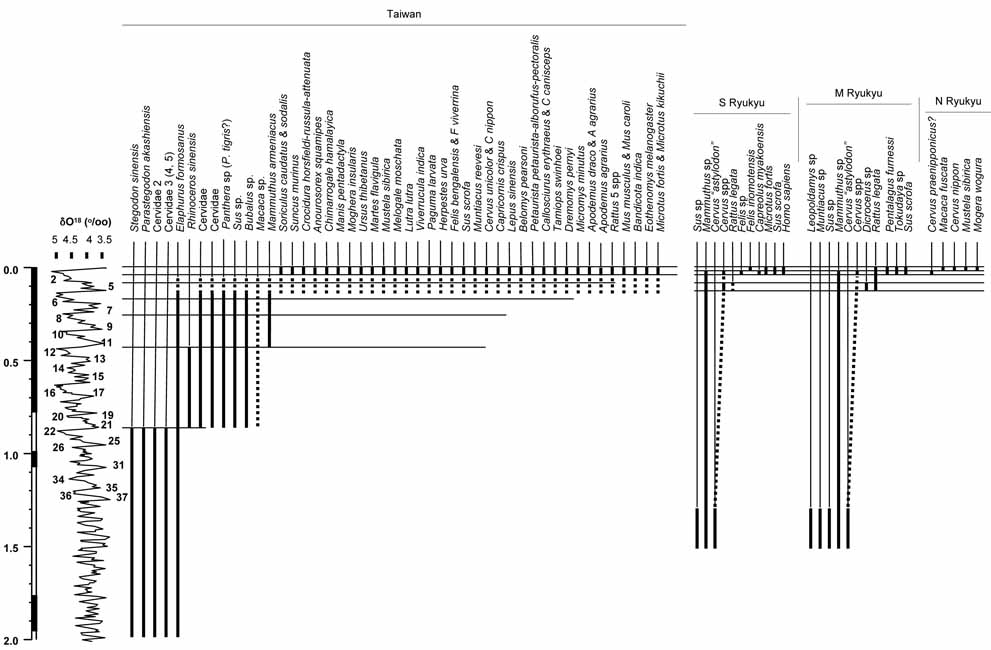
Range chart of the Quaternary mammals from Taiwan and the Ryukyu Islands (Van der Made, 2005).
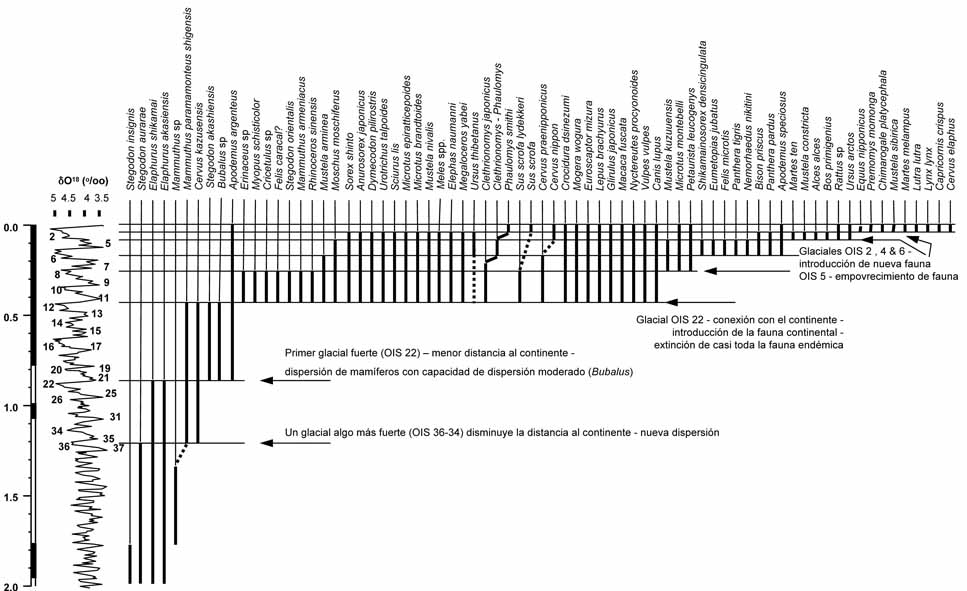
Range chart of the Quaternary mammals from Japan (Kyushu and Honshu) (Van der Made, 2005).
- Made, J. van der, 2005. La fauna del Pleistoceno europeo. In: E. Carbonell, X.P. Rodríguez, R. Sala, J. van der Made, C. Lorenzo, M. Mosquera, M. Vaquero, J. Rosell, J. Vallverdú, F. Burjachs, P. Hortolà. Homínidos: las primeras ocupaciones de los continentes. Capítulo 4 - Europa; Sección 4.4. Ariel: 394-432.
- Made, J. van der, 1999. Biogeography and stratigraphy of the Mio-Pleistocene mammals of Sardinia and the description of some fossils. Deinsea, 7: 337-360.
- Made, J. van der, 1988. Sus nanus nov. sp. a Pliocene dwarf pig from Capo Figari (Sardinia). Bolletino della Società Paleontologica Italiana, 27 (3): 367-378, 3 pls.
5) Others
- Zliobaité, I., M. Fortelius, R.L. Bernor, L.W. van den Hoek Ostende, C.M. Janis, K. Lintulaakso, L.K. Saila, L. Werdelin, I. Casanovas-Vilar, D.A. Croft, L.J. Flynn, S.S.B. Hopkins, A. Kaakinen, L. Kordos, D.S. Kostopoulos, L. Pandolfi, J. Rowan, A. Tesakov, I. Vislobokova, Z. Zhang, M. Aiglstorfer, D.M. Alba, M. Arnal, P.-O. Antoine, M. Belmaker, M. Bilgin, J.-R. Boisserie, M.R. Borths, S.B. Cooke, J.A. van Dam, E. Delson, J.T. Eronen, D. Fox, A.R. Friscia, M. Furio, I.X. Giaourtsakis, L. Holbrook, J. Hunter, S. Lopez-Torres, J. Ludtke, R. Minwer-Barakat, J. van der Made, B. Mennecart, D. Pushkina, L. Rook, J. Saarinen, J.X. Samuels, W. Sanders, M.T. Silcox & J. Vepsalainen, 2023. The NOW Database of Fossil Mammals. In: I. Casanovas-Vilar, L.W. van den Hoek Ostende, C.M. Janis, J. Saarinen (eds.), Evolution of Cenozoic Land Mammal Faunas and Ecosystems: 25 Years of the NOW Database of Fossil Mammals. Vertebrate Paleobiology and Paleoanthropology series. Springer: 33-42.
I am involved with the NOW database since the meeting in Reisensburg in 1992 and am coordinator for the Suoidea.
- Made, J. van der, 2023. El campo no es un vertedero. Naturalmente, 37: 29-33.
- Made, J. van der, 2004. Das Pleistozän von Untermassfeld bei Meiningen (Thüringen) Teil 2 & Teil 3. Geobios, 37/4: 553-554.
Otros investigadores del departamento
- Fernández Jalvo Yolanda
- Alberdi Alonso María Teresa
- Bastir Markus
- Bruner Emiliano
- Caballero Chordá Óscar
- Cantalapiedra Juan L
- Darío Fidalgo Casares
- Estalrrich Albo Almudena
- Fernandez Cascon Beatriz
- Garcia Martinez Daniel
- Garcia Morato Sara
- García Tabernero Antonio
- Gómez Recio Marta
- Gutiérrez Aida
- Knoll Fabien
- López Cano Miguel
- López-Rey José M.
- Macho-Callejo Alba
- Madroñal López Gonzalo
- Martín Perea David Manuel
- Morales Romero Jorge
- Palancar Carlos A.
- Pelaez-Campomanes de_Labra Pablo
- Rodrigo Córdoba Clara
- Rosas Antonio
- Salesa Calvo Manuel Jesús
- Sanz Prieto Daniel
- Siliceo Gema
- Torres-Medina Tomás